生物吸附法去除重金属离子的研究进展
1
2010
... 吸附技术是简单、高效去除废水中污染物的方法之一.重金属微生物吸附是以细菌、真菌和藻类等微生物为吸附剂吸附液相中的重金属离子,之后再通过固液分离去除重金属离子的一种方法〔1〕.根据微生物细胞的不同活性,微生物吸附重金属可分为活/生长细胞吸附和死细胞吸附2种方法.尽管二者各有优缺点,但利用微生物活细胞吸附重金属是未来生物吸附领域的研究方向〔2-3〕.然而,到目前为止,重金属废水的微生物活细胞处理技术并没有在实际废水处理中得到广泛应用,其大多停留在实验室或中试研究水平〔4〕,导致这种情况的重要原因之一就是重金属对微生物具有毒性和生长抑制作用. ...
Research advances in heavy metal removal by biosorption
1
2010
... 吸附技术是简单、高效去除废水中污染物的方法之一.重金属微生物吸附是以细菌、真菌和藻类等微生物为吸附剂吸附液相中的重金属离子,之后再通过固液分离去除重金属离子的一种方法〔1〕.根据微生物细胞的不同活性,微生物吸附重金属可分为活/生长细胞吸附和死细胞吸附2种方法.尽管二者各有优缺点,但利用微生物活细胞吸附重金属是未来生物吸附领域的研究方向〔2-3〕.然而,到目前为止,重金属废水的微生物活细胞处理技术并没有在实际废水处理中得到广泛应用,其大多停留在实验室或中试研究水平〔4〕,导致这种情况的重要原因之一就是重金属对微生物具有毒性和生长抑制作用. ...
Metal bioremediation through growing cells
1
2004
... 吸附技术是简单、高效去除废水中污染物的方法之一.重金属微生物吸附是以细菌、真菌和藻类等微生物为吸附剂吸附液相中的重金属离子,之后再通过固液分离去除重金属离子的一种方法〔1〕.根据微生物细胞的不同活性,微生物吸附重金属可分为活/生长细胞吸附和死细胞吸附2种方法.尽管二者各有优缺点,但利用微生物活细胞吸附重金属是未来生物吸附领域的研究方向〔2-3〕.然而,到目前为止,重金属废水的微生物活细胞处理技术并没有在实际废水处理中得到广泛应用,其大多停留在实验室或中试研究水平〔4〕,导致这种情况的重要原因之一就是重金属对微生物具有毒性和生长抑制作用. ...
Biosorption:Critical review of scientific rationale,environmental importance and significance for pollution treatment
1
2009
... 吸附技术是简单、高效去除废水中污染物的方法之一.重金属微生物吸附是以细菌、真菌和藻类等微生物为吸附剂吸附液相中的重金属离子,之后再通过固液分离去除重金属离子的一种方法〔1〕.根据微生物细胞的不同活性,微生物吸附重金属可分为活/生长细胞吸附和死细胞吸附2种方法.尽管二者各有优缺点,但利用微生物活细胞吸附重金属是未来生物吸附领域的研究方向〔2-3〕.然而,到目前为止,重金属废水的微生物活细胞处理技术并没有在实际废水处理中得到广泛应用,其大多停留在实验室或中试研究水平〔4〕,导致这种情况的重要原因之一就是重金属对微生物具有毒性和生长抑制作用. ...
1
2015
... 吸附技术是简单、高效去除废水中污染物的方法之一.重金属微生物吸附是以细菌、真菌和藻类等微生物为吸附剂吸附液相中的重金属离子,之后再通过固液分离去除重金属离子的一种方法〔1〕.根据微生物细胞的不同活性,微生物吸附重金属可分为活/生长细胞吸附和死细胞吸附2种方法.尽管二者各有优缺点,但利用微生物活细胞吸附重金属是未来生物吸附领域的研究方向〔2-3〕.然而,到目前为止,重金属废水的微生物活细胞处理技术并没有在实际废水处理中得到广泛应用,其大多停留在实验室或中试研究水平〔4〕,导致这种情况的重要原因之一就是重金属对微生物具有毒性和生长抑制作用. ...
1
2015
... 吸附技术是简单、高效去除废水中污染物的方法之一.重金属微生物吸附是以细菌、真菌和藻类等微生物为吸附剂吸附液相中的重金属离子,之后再通过固液分离去除重金属离子的一种方法〔1〕.根据微生物细胞的不同活性,微生物吸附重金属可分为活/生长细胞吸附和死细胞吸附2种方法.尽管二者各有优缺点,但利用微生物活细胞吸附重金属是未来生物吸附领域的研究方向〔2-3〕.然而,到目前为止,重金属废水的微生物活细胞处理技术并没有在实际废水处理中得到广泛应用,其大多停留在实验室或中试研究水平〔4〕,导致这种情况的重要原因之一就是重金属对微生物具有毒性和生长抑制作用. ...
Multiple transcriptional mechanisms collectively mediate copper resistance in Cupriavidus gilardii CR3
2
2019
... 如何突破重金属废水微生物处理的技术瓶颈一直是工业水处理领域的研究热点.重金属抗性微生物是一类可以在重金属胁迫下生长,比通常微生物具有更高重金属抵抗能力的微生物,极具理论研究价值和应用价值,近年来在环境污染生物修复领域受到关注和重视〔5〕.但是,目前人们对重金属抗性微生物的特性和应用仍然知之甚少.笔者分别以“Metal biosorption”、“Metal biosorption and resistant”为检索主题词在Web of Science数据库进行检索,结果见表1. ...
... 细菌对重金属的生物累积存在2种可能机制.一种是重金属离子通过细胞膜进入到细胞质中,在细胞内特定的区域被隔离,以防止其接触到重要的细胞组分或细胞器〔41〕,另一种是重金属离子与细菌体内有生物活性的配位体发生特异的生物化学反应后形成螯合物对细菌进行生物解毒〔42〕.原核生物(如细菌)胞内含有谷胱甘肽(Glutathione,GSH)、金属硫蛋白等,这些物质可与重金属离子结合形成螯合物,对于重金属的解毒和防御重金属引起的氧化胁迫具有重要作用.GSH是由谷氨酸、半胱氨酸及甘氨酸组成的三肽,其含有的氨基、巯基、羧基和酰胺基等多种配位基团可与重金属离子结合,在生物体内对重金属进行累积.GSH还可以清除因重金属胁迫产生的活性氧自由基,以此来降低重金属离子对细胞的生物毒性〔43〕.细菌的重金属抗性机制是菌细胞应对重金属胁迫产生的一系列复杂的生理生化反应,Ning HUANG等〔5〕通过基因组与转录组学研究表明细菌对重金属的抗性往往不依赖于单一的机制,而是多种系统相互作用的结果. ...
Multimetal resistance and tolerance in microbial biofilms
1
2007
... 作为一种开放的生命体,微生物在其生长过程中不可避免地受到极端环境条件(温度、pH、干旱)和重金属胁迫等影响.当栖息在重金属环境时,部分微生物被发现具有重金属抗性.重金属抗性是指微生物在多种有毒重(准)金属离子存在下能够连续生长的能力〔6〕.重金属抗性微生物是某些可以在重金属胁迫条件下连续生长的特定微生物的总称,包括重金属抗性细菌、微藻和真菌等.相对而言,对重金属抗性微生物的研究目前多集中在对重金属抗性细菌的研究上〔7〕. ...
Metal resistance and its association with antibiotic resistance
1
2017
... 作为一种开放的生命体,微生物在其生长过程中不可避免地受到极端环境条件(温度、pH、干旱)和重金属胁迫等影响.当栖息在重金属环境时,部分微生物被发现具有重金属抗性.重金属抗性是指微生物在多种有毒重(准)金属离子存在下能够连续生长的能力〔6〕.重金属抗性微生物是某些可以在重金属胁迫条件下连续生长的特定微生物的总称,包括重金属抗性细菌、微藻和真菌等.相对而言,对重金属抗性微生物的研究目前多集中在对重金属抗性细菌的研究上〔7〕. ...
tcrB,a gene conferring transferable copper resistance in Enterococcus faecium:Occurrence,transferability,and linkage to macrolide and glycopeptide resistance
1
2002
... 对重金属抗性细菌的研究最早可追溯到20世纪60、70年代,其主要与硫酸铜在养猪和农业种植中的使用有关.硫酸铜可以使猪肠道内微生物菌群发生变化〔8〕,因此被用作饲料添加剂以使猪的体重增加,研究者在以硫酸铜作为饲料添加剂的养猪场废水中分离出具有铜抗性的Escherichia coli〔9-10〕.作为金属无机盐,硫酸铜还常被用作农作物病原菌的杀菌剂,其长期使用后,杀菌效果降低,原因是某些植物病原菌对铜离子产生了抗性,此外,研究人员还在喷洒硫酸铜作为杀菌剂的作物如烟草、土豆、番茄、胡椒、猕猴桃受病原菌侵扰的叶片上分离出多种重金属铜的抗性菌,如Xanthomonas campestris pathovar(pv.) vesicatoria、Pseudomonas syringae pv. tomato〔11-12〕、Escherichia coli〔13-14〕和Mycobacterium scrofulaceum〔15〕. ...
Resistogram typing:A new epidemiological tool—Application to Escherichia coli
1
1970
... 对重金属抗性细菌的研究最早可追溯到20世纪60、70年代,其主要与硫酸铜在养猪和农业种植中的使用有关.硫酸铜可以使猪肠道内微生物菌群发生变化〔8〕,因此被用作饲料添加剂以使猪的体重增加,研究者在以硫酸铜作为饲料添加剂的养猪场废水中分离出具有铜抗性的Escherichia coli〔9-10〕.作为金属无机盐,硫酸铜还常被用作农作物病原菌的杀菌剂,其长期使用后,杀菌效果降低,原因是某些植物病原菌对铜离子产生了抗性,此外,研究人员还在喷洒硫酸铜作为杀菌剂的作物如烟草、土豆、番茄、胡椒、猕猴桃受病原菌侵扰的叶片上分离出多种重金属铜的抗性菌,如Xanthomonas campestris pathovar(pv.) vesicatoria、Pseudomonas syringae pv. tomato〔11-12〕、Escherichia coli〔13-14〕和Mycobacterium scrofulaceum〔15〕. ...
Resistance transfer fecal coliforms isolated from the Whippany river
1
1974
... 对重金属抗性细菌的研究最早可追溯到20世纪60、70年代,其主要与硫酸铜在养猪和农业种植中的使用有关.硫酸铜可以使猪肠道内微生物菌群发生变化〔8〕,因此被用作饲料添加剂以使猪的体重增加,研究者在以硫酸铜作为饲料添加剂的养猪场废水中分离出具有铜抗性的Escherichia coli〔9-10〕.作为金属无机盐,硫酸铜还常被用作农作物病原菌的杀菌剂,其长期使用后,杀菌效果降低,原因是某些植物病原菌对铜离子产生了抗性,此外,研究人员还在喷洒硫酸铜作为杀菌剂的作物如烟草、土豆、番茄、胡椒、猕猴桃受病原菌侵扰的叶片上分离出多种重金属铜的抗性菌,如Xanthomonas campestris pathovar(pv.) vesicatoria、Pseudomonas syringae pv. tomato〔11-12〕、Escherichia coli〔13-14〕和Mycobacterium scrofulaceum〔15〕. ...
Copper-tolerance in Pseudomonas syringae pv. tomato and Xanthomonas spp. and the control of diseases associated with these pathogens in tomato and pepper. A systematic literature review
1
2017
... 对重金属抗性细菌的研究最早可追溯到20世纪60、70年代,其主要与硫酸铜在养猪和农业种植中的使用有关.硫酸铜可以使猪肠道内微生物菌群发生变化〔8〕,因此被用作饲料添加剂以使猪的体重增加,研究者在以硫酸铜作为饲料添加剂的养猪场废水中分离出具有铜抗性的Escherichia coli〔9-10〕.作为金属无机盐,硫酸铜还常被用作农作物病原菌的杀菌剂,其长期使用后,杀菌效果降低,原因是某些植物病原菌对铜离子产生了抗性,此外,研究人员还在喷洒硫酸铜作为杀菌剂的作物如烟草、土豆、番茄、胡椒、猕猴桃受病原菌侵扰的叶片上分离出多种重金属铜的抗性菌,如Xanthomonas campestris pathovar(pv.) vesicatoria、Pseudomonas syringae pv. tomato〔11-12〕、Escherichia coli〔13-14〕和Mycobacterium scrofulaceum〔15〕. ...
Similarity between copper resistance genes from Pseudomonas syringae pv. actinidiae and P. syringae pv. tomato
1
2002
... 对重金属抗性细菌的研究最早可追溯到20世纪60、70年代,其主要与硫酸铜在养猪和农业种植中的使用有关.硫酸铜可以使猪肠道内微生物菌群发生变化〔8〕,因此被用作饲料添加剂以使猪的体重增加,研究者在以硫酸铜作为饲料添加剂的养猪场废水中分离出具有铜抗性的Escherichia coli〔9-10〕.作为金属无机盐,硫酸铜还常被用作农作物病原菌的杀菌剂,其长期使用后,杀菌效果降低,原因是某些植物病原菌对铜离子产生了抗性,此外,研究人员还在喷洒硫酸铜作为杀菌剂的作物如烟草、土豆、番茄、胡椒、猕猴桃受病原菌侵扰的叶片上分离出多种重金属铜的抗性菌,如Xanthomonas campestris pathovar(pv.) vesicatoria、Pseudomonas syringae pv. tomato〔11-12〕、Escherichia coli〔13-14〕和Mycobacterium scrofulaceum〔15〕. ...
Inducible plasmid-mediated copper resistance in Escherichia coli
1
1985
... 对重金属抗性细菌的研究最早可追溯到20世纪60、70年代,其主要与硫酸铜在养猪和农业种植中的使用有关.硫酸铜可以使猪肠道内微生物菌群发生变化〔8〕,因此被用作饲料添加剂以使猪的体重增加,研究者在以硫酸铜作为饲料添加剂的养猪场废水中分离出具有铜抗性的Escherichia coli〔9-10〕.作为金属无机盐,硫酸铜还常被用作农作物病原菌的杀菌剂,其长期使用后,杀菌效果降低,原因是某些植物病原菌对铜离子产生了抗性,此外,研究人员还在喷洒硫酸铜作为杀菌剂的作物如烟草、土豆、番茄、胡椒、猕猴桃受病原菌侵扰的叶片上分离出多种重金属铜的抗性菌,如Xanthomonas campestris pathovar(pv.) vesicatoria、Pseudomonas syringae pv. tomato〔11-12〕、Escherichia coli〔13-14〕和Mycobacterium scrofulaceum〔15〕. ...
Plasmid-controlled resistance to copper in Escherichia coli
1
1983
... 对重金属抗性细菌的研究最早可追溯到20世纪60、70年代,其主要与硫酸铜在养猪和农业种植中的使用有关.硫酸铜可以使猪肠道内微生物菌群发生变化〔8〕,因此被用作饲料添加剂以使猪的体重增加,研究者在以硫酸铜作为饲料添加剂的养猪场废水中分离出具有铜抗性的Escherichia coli〔9-10〕.作为金属无机盐,硫酸铜还常被用作农作物病原菌的杀菌剂,其长期使用后,杀菌效果降低,原因是某些植物病原菌对铜离子产生了抗性,此外,研究人员还在喷洒硫酸铜作为杀菌剂的作物如烟草、土豆、番茄、胡椒、猕猴桃受病原菌侵扰的叶片上分离出多种重金属铜的抗性菌,如Xanthomonas campestris pathovar(pv.) vesicatoria、Pseudomonas syringae pv. tomato〔11-12〕、Escherichia coli〔13-14〕和Mycobacterium scrofulaceum〔15〕. ...
Plasmid-encoded copper resistance and precipitation by Mycobacterium scrofulaceum
1
1987
... 对重金属抗性细菌的研究最早可追溯到20世纪60、70年代,其主要与硫酸铜在养猪和农业种植中的使用有关.硫酸铜可以使猪肠道内微生物菌群发生变化〔8〕,因此被用作饲料添加剂以使猪的体重增加,研究者在以硫酸铜作为饲料添加剂的养猪场废水中分离出具有铜抗性的Escherichia coli〔9-10〕.作为金属无机盐,硫酸铜还常被用作农作物病原菌的杀菌剂,其长期使用后,杀菌效果降低,原因是某些植物病原菌对铜离子产生了抗性,此外,研究人员还在喷洒硫酸铜作为杀菌剂的作物如烟草、土豆、番茄、胡椒、猕猴桃受病原菌侵扰的叶片上分离出多种重金属铜的抗性菌,如Xanthomonas campestris pathovar(pv.) vesicatoria、Pseudomonas syringae pv. tomato〔11-12〕、Escherichia coli〔13-14〕和Mycobacterium scrofulaceum〔15〕. ...
Resistance to inorganic salts and antibiotics among sewage-borne Enterobacteriaceae and Achromobacteriaceae
1
1976
... 由于细菌对重金属与抗生素具有协同抗性和交叉抗性,临床医学中也涉及到重金属抗性菌的分离,如银作为杀菌药物用于临床医学,研究人员在伤口附近分离出重金属银的抗性菌.在重金属抗性细菌研究早期,已报道的重金属抗性菌主要为肠杆菌科(Enterobacteriaceae)、假单胞菌科(Pseudomonadaceae)、无色细菌科(Achromobacteriaceae)和螺旋菌科(Spirillaceae)细菌等〔16〕. ...
Extrachromosomal inheritance controlling resistance to cadmium,cobalt,copper and zinc ions:Evidence from curing in a Pseudomonas proceedings
1
1978
... 自20世纪70年代后,不同种类的重金属抗性细菌陆续不断地被分离出来,为微生物重金属抗性机制的研究与应用奠定了基础.丁香假单胞菌、大肠杆菌和海氏肠球菌是重金属抗性细菌抗性机制研究的模式菌.此外,贪铜菌属细菌(Cupriavidus)在重金属抗性机制研究中占有重要的地位,是目前已有报道的重金属抗性细菌中比较特殊的一类.贪铜菌属细菌分布广泛,在比利时、美国、中国、德国、新西兰和日本等地都有检出.该菌属最显著的特征是其对铜离子具有抗性,铜离子对贪铜菌属细菌的细胞繁殖没有影响,甚至可能促进其生长.贪铜菌属细菌Cupriavidus metallidurans CH34是重金属抗性外排系统研究的典型代表菌种,该菌株在比利时炼锌厂滗析槽污泥中被分离获得.由于滗析槽中含有较高浓度的重金属锌、镉和铜等〔17-18〕,菌株CH34对Cd、Co、Cu、Hg、Ni、Pb、Tl、Zn、Cr等多种重金属具有抗性作用.菌株CH34的抗性基因位于pMOL28(171 kb)和pMOL30(234 kb)2个大质粒上,质粒pMOL28携带铬、镍、钴抗性基因,质粒pMOL30携带锌、钴、铅、铜抗性基因〔19〕. ...
Ralstonia metallidurans,a bacterium specifically adapted to toxic metals:Towards a catalogue of metal-responsive genes
1
2003
... 自20世纪70年代后,不同种类的重金属抗性细菌陆续不断地被分离出来,为微生物重金属抗性机制的研究与应用奠定了基础.丁香假单胞菌、大肠杆菌和海氏肠球菌是重金属抗性细菌抗性机制研究的模式菌.此外,贪铜菌属细菌(Cupriavidus)在重金属抗性机制研究中占有重要的地位,是目前已有报道的重金属抗性细菌中比较特殊的一类.贪铜菌属细菌分布广泛,在比利时、美国、中国、德国、新西兰和日本等地都有检出.该菌属最显著的特征是其对铜离子具有抗性,铜离子对贪铜菌属细菌的细胞繁殖没有影响,甚至可能促进其生长.贪铜菌属细菌Cupriavidus metallidurans CH34是重金属抗性外排系统研究的典型代表菌种,该菌株在比利时炼锌厂滗析槽污泥中被分离获得.由于滗析槽中含有较高浓度的重金属锌、镉和铜等〔17-18〕,菌株CH34对Cd、Co、Cu、Hg、Ni、Pb、Tl、Zn、Cr等多种重金属具有抗性作用.菌株CH34的抗性基因位于pMOL28(171 kb)和pMOL30(234 kb)2个大质粒上,质粒pMOL28携带铬、镍、钴抗性基因,质粒pMOL30携带锌、钴、铅、铜抗性基因〔19〕. ...
Cupriavidus metallidurans:Evolution of a metal-resistant bacterium
1
2009
... 自20世纪70年代后,不同种类的重金属抗性细菌陆续不断地被分离出来,为微生物重金属抗性机制的研究与应用奠定了基础.丁香假单胞菌、大肠杆菌和海氏肠球菌是重金属抗性细菌抗性机制研究的模式菌.此外,贪铜菌属细菌(Cupriavidus)在重金属抗性机制研究中占有重要的地位,是目前已有报道的重金属抗性细菌中比较特殊的一类.贪铜菌属细菌分布广泛,在比利时、美国、中国、德国、新西兰和日本等地都有检出.该菌属最显著的特征是其对铜离子具有抗性,铜离子对贪铜菌属细菌的细胞繁殖没有影响,甚至可能促进其生长.贪铜菌属细菌Cupriavidus metallidurans CH34是重金属抗性外排系统研究的典型代表菌种,该菌株在比利时炼锌厂滗析槽污泥中被分离获得.由于滗析槽中含有较高浓度的重金属锌、镉和铜等〔17-18〕,菌株CH34对Cd、Co、Cu、Hg、Ni、Pb、Tl、Zn、Cr等多种重金属具有抗性作用.菌株CH34的抗性基因位于pMOL28(171 kb)和pMOL30(234 kb)2个大质粒上,质粒pMOL28携带铬、镍、钴抗性基因,质粒pMOL30携带锌、钴、铅、铜抗性基因〔19〕. ...
Determination of minimum inhibitory concentrations
1
2001
... 在重金属抗性细菌研究过程中,如何判断某种细菌对重金属具有抗性是一个首要问题.大多数研究采用最小抑菌浓度(Minimum inhibitory concentration,MIC)评价细菌对某种抑菌剂(抗生素、重金属离子或杀菌剂)的抗性能力〔20〕.为了确定细菌对某种重金属的MIC,通常在细菌培养基中添加不同剂量的重金属盐,能完全抑制细菌生长的该重金属离子的最小浓度即为MIC.尽管MIC性能测试实验可能由于重金属离子与细菌培养基的某种成分产生螯合作用导致重金属离子浓度与实际浓度不完全相符,但其仍然是评估细菌对重金属抗性作用的一种有效方法〔21〕. ...
Effects of heavy metals on Pseudomonas aeruginosa and Bacillus thuringiensis
2
1998
... 在重金属抗性细菌研究过程中,如何判断某种细菌对重金属具有抗性是一个首要问题.大多数研究采用最小抑菌浓度(Minimum inhibitory concentration,MIC)评价细菌对某种抑菌剂(抗生素、重金属离子或杀菌剂)的抗性能力〔20〕.为了确定细菌对某种重金属的MIC,通常在细菌培养基中添加不同剂量的重金属盐,能完全抑制细菌生长的该重金属离子的最小浓度即为MIC.尽管MIC性能测试实验可能由于重金属离子与细菌培养基的某种成分产生螯合作用导致重金属离子浓度与实际浓度不完全相符,但其仍然是评估细菌对重金属抗性作用的一种有效方法〔21〕. ...
... Heavy metal resistant bacteria and their minimum inhibitory concentration(MIC)
Table 2| 细菌 | 编号 | MIC/(mmol·L-1) | 参考文献 |
|---|
| Cd | Co | Cu | Zn | Hg |
|---|
| Bacillus thuringiensis | / | 1.2 | 0.05 | 0.5 | 0.5 | 0.06 | 〔21〕 |
| Cupriavidus metallidurans | CH34 | 4 | 25 | 3 | 12 | 0.002 7 | 〔22〕 |
| Cupriavidus gilardii | CR3 | 4 | 2 | 3 | >24 | >0.04 | 〔23〕 |
| Citrobacterfreundii | JPG1 | — | — | 2 | — | — | 〔24〕 |
| Pseudomonas aeruginosa | S7 | 1.5 | 0.6 | 1.6 | 1.5 | 0.05 | 〔25〕 |
| Pseudomonas cepacia | S14 | 0.2 | 0.2 | 1.2 | 0.2 | 0.04 | 〔25〕 |
| Pseudomonas paucimobiliz | S12 | 0.2 | 0.2 | 1.5 | 0.5 | 0.05 | 〔25〕 |
| Pseudomonas putida | CZ1 | — | — | 5 | 5 | — | 〔26〕 |
注:—表示无测定值. ...
Heavy metal resistance in Cupriavidus metallidurans CH34 is governed by an intricate transcriptional network
1
2011
... Heavy metal resistant bacteria and their minimum inhibitory concentration(MIC)
Table 2| 细菌 | 编号 | MIC/(mmol·L-1) | 参考文献 |
|---|
| Cd | Co | Cu | Zn | Hg |
|---|
| Bacillus thuringiensis | / | 1.2 | 0.05 | 0.5 | 0.5 | 0.06 | 〔21〕 |
| Cupriavidus metallidurans | CH34 | 4 | 25 | 3 | 12 | 0.002 7 | 〔22〕 |
| Cupriavidus gilardii | CR3 | 4 | 2 | 3 | >24 | >0.04 | 〔23〕 |
| Citrobacterfreundii | JPG1 | — | — | 2 | — | — | 〔24〕 |
| Pseudomonas aeruginosa | S7 | 1.5 | 0.6 | 1.6 | 1.5 | 0.05 | 〔25〕 |
| Pseudomonas cepacia | S14 | 0.2 | 0.2 | 1.2 | 0.2 | 0.04 | 〔25〕 |
| Pseudomonas paucimobiliz | S12 | 0.2 | 0.2 | 1.5 | 0.5 | 0.05 | 〔25〕 |
| Pseudomonas putida | CZ1 | — | — | 5 | 5 | — | 〔26〕 |
注:—表示无测定值. ...
Genome sequence analysis of the naphthenic acid degrading and metal resistant bacterium Cupriavidus gilardii CR3
1
2015
... Heavy metal resistant bacteria and their minimum inhibitory concentration(MIC)
Table 2| 细菌 | 编号 | MIC/(mmol·L-1) | 参考文献 |
|---|
| Cd | Co | Cu | Zn | Hg |
|---|
| Bacillus thuringiensis | / | 1.2 | 0.05 | 0.5 | 0.5 | 0.06 | 〔21〕 |
| Cupriavidus metallidurans | CH34 | 4 | 25 | 3 | 12 | 0.002 7 | 〔22〕 |
| Cupriavidus gilardii | CR3 | 4 | 2 | 3 | >24 | >0.04 | 〔23〕 |
| Citrobacterfreundii | JPG1 | — | — | 2 | — | — | 〔24〕 |
| Pseudomonas aeruginosa | S7 | 1.5 | 0.6 | 1.6 | 1.5 | 0.05 | 〔25〕 |
| Pseudomonas cepacia | S14 | 0.2 | 0.2 | 1.2 | 0.2 | 0.04 | 〔25〕 |
| Pseudomonas paucimobiliz | S12 | 0.2 | 0.2 | 1.5 | 0.5 | 0.05 | 〔25〕 |
| Pseudomonas putida | CZ1 | — | — | 5 | 5 | — | 〔26〕 |
注:—表示无测定值. ...
Coupling heavy metal resistance and oxygen flexibility for bioremoval of copper ions by newly isolated Citrobacter freundii JPG1
1
2018
... Heavy metal resistant bacteria and their minimum inhibitory concentration(MIC)
Table 2| 细菌 | 编号 | MIC/(mmol·L-1) | 参考文献 |
|---|
| Cd | Co | Cu | Zn | Hg |
|---|
| Bacillus thuringiensis | / | 1.2 | 0.05 | 0.5 | 0.5 | 0.06 | 〔21〕 |
| Cupriavidus metallidurans | CH34 | 4 | 25 | 3 | 12 | 0.002 7 | 〔22〕 |
| Cupriavidus gilardii | CR3 | 4 | 2 | 3 | >24 | >0.04 | 〔23〕 |
| Citrobacterfreundii | JPG1 | — | — | 2 | — | — | 〔24〕 |
| Pseudomonas aeruginosa | S7 | 1.5 | 0.6 | 1.6 | 1.5 | 0.05 | 〔25〕 |
| Pseudomonas cepacia | S14 | 0.2 | 0.2 | 1.2 | 0.2 | 0.04 | 〔25〕 |
| Pseudomonas paucimobiliz | S12 | 0.2 | 0.2 | 1.5 | 0.5 | 0.05 | 〔25〕 |
| Pseudomonas putida | CZ1 | — | — | 5 | 5 | — | 〔26〕 |
注:—表示无测定值. ...
Resistance of environmental bacteria to heavy metals
3
1998
... Heavy metal resistant bacteria and their minimum inhibitory concentration(MIC)
Table 2| 细菌 | 编号 | MIC/(mmol·L-1) | 参考文献 |
|---|
| Cd | Co | Cu | Zn | Hg |
|---|
| Bacillus thuringiensis | / | 1.2 | 0.05 | 0.5 | 0.5 | 0.06 | 〔21〕 |
| Cupriavidus metallidurans | CH34 | 4 | 25 | 3 | 12 | 0.002 7 | 〔22〕 |
| Cupriavidus gilardii | CR3 | 4 | 2 | 3 | >24 | >0.04 | 〔23〕 |
| Citrobacterfreundii | JPG1 | — | — | 2 | — | — | 〔24〕 |
| Pseudomonas aeruginosa | S7 | 1.5 | 0.6 | 1.6 | 1.5 | 0.05 | 〔25〕 |
| Pseudomonas cepacia | S14 | 0.2 | 0.2 | 1.2 | 0.2 | 0.04 | 〔25〕 |
| Pseudomonas paucimobiliz | S12 | 0.2 | 0.2 | 1.5 | 0.5 | 0.05 | 〔25〕 |
| Pseudomonas putida | CZ1 | — | — | 5 | 5 | — | 〔26〕 |
注:—表示无测定值. ...
... 〔
25〕
| Pseudomonas paucimobiliz | S12 | 0.2 | 0.2 | 1.5 | 0.5 | 0.05 | 〔25〕 |
| Pseudomonas putida | CZ1 | — | — | 5 | 5 | — | 〔26〕 |
注:—表示无测定值. ...
... 〔
25〕
| Pseudomonas putida | CZ1 | — | — | 5 | 5 | — | 〔26〕 |
注:—表示无测定值. ...
Tolerance and biosorption of copper and zinc by Pseudomonas putida CZ1 isolated from metal-polluted soil
2
2006
... Heavy metal resistant bacteria and their minimum inhibitory concentration(MIC)
Table 2| 细菌 | 编号 | MIC/(mmol·L-1) | 参考文献 |
|---|
| Cd | Co | Cu | Zn | Hg |
|---|
| Bacillus thuringiensis | / | 1.2 | 0.05 | 0.5 | 0.5 | 0.06 | 〔21〕 |
| Cupriavidus metallidurans | CH34 | 4 | 25 | 3 | 12 | 0.002 7 | 〔22〕 |
| Cupriavidus gilardii | CR3 | 4 | 2 | 3 | >24 | >0.04 | 〔23〕 |
| Citrobacterfreundii | JPG1 | — | — | 2 | — | — | 〔24〕 |
| Pseudomonas aeruginosa | S7 | 1.5 | 0.6 | 1.6 | 1.5 | 0.05 | 〔25〕 |
| Pseudomonas cepacia | S14 | 0.2 | 0.2 | 1.2 | 0.2 | 0.04 | 〔25〕 |
| Pseudomonas paucimobiliz | S12 | 0.2 | 0.2 | 1.5 | 0.5 | 0.05 | 〔25〕 |
| Pseudomonas putida | CZ1 | — | — | 5 | 5 | — | 〔26〕 |
注:—表示无测定值. ...
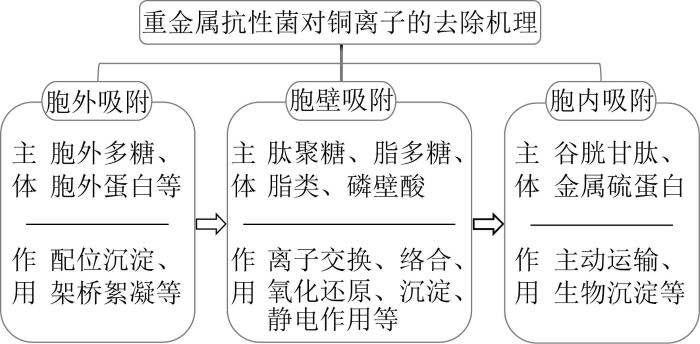
... EPS中的大分子含有多种可以与重金属离子结合的阴离子基团,如羧基、羟基、巯基和磷酸根基团等,各阴离子基团通过配位、吸附、沉淀和架桥絮凝等作用吸附重金属离子,防止重金属离子与细胞作用〔37〕.研究发现适当的重金属离子浓度可以诱导细菌增加EPS的分泌,即在一定浓度的重金属环境中,细菌对重金属胁迫产生响应而上调胞外聚合物基因的表达,从而增加胞外聚合物对重金属离子的捕获,如Pseudomonas putida〔26〕、Pseudomonas aeruginosa〔38〕、B. subtilis〔39〕在铜的作用下,EPS都有明显的增加.反之,如果去除菌属细菌产生的胞外多糖,细菌吸附重金属能力明显降低,对重金属的敏感性增加. ...
Microbial resistance to metals in the environment
2
2000
... 为了避免过量重金属离子对细胞的生物毒性,细菌在长期的进化过程中形成了复杂多样的重金属抗性机制以维持正常的细胞内重金属离子浓度.依据不同的角度和理解,研究者对细菌的重金属抗性机制有着不同的划分〔27-28〕,如R. M. MAIER等〔28〕认为重金属抗性机制包括一般抗性机制和依赖金属的特殊抗性机制;J. A. LEMIRE等〔29〕把细菌的重金属抗性机制归纳为如图1所示的5个方面,包括细胞外隔离、减少摄取、外排系统、胞内累积和解毒机制,其认为当环境中存在过量重金属时,细菌可通过渗透屏障、胞外蛋白/螯合剂与被动容忍作用减少对重金属离子的摄取,当重金属离子跨膜进入胞内后,细菌可通过外排系统将胞内重金属离子主动运输到胞外.此外,细菌还可通过重金属与胞内蛋白/螯合剂结合、酶解毒、降低重金属靶标敏感性等多重作用使重金属在胞内进行累积和解毒〔27,30〕. ...
... 〔27,30〕. ...
2
2004
... 为了避免过量重金属离子对细胞的生物毒性,细菌在长期的进化过程中形成了复杂多样的重金属抗性机制以维持正常的细胞内重金属离子浓度.依据不同的角度和理解,研究者对细菌的重金属抗性机制有着不同的划分〔27-28〕,如R. M. MAIER等〔28〕认为重金属抗性机制包括一般抗性机制和依赖金属的特殊抗性机制;J. A. LEMIRE等〔29〕把细菌的重金属抗性机制归纳为如图1所示的5个方面,包括细胞外隔离、减少摄取、外排系统、胞内累积和解毒机制,其认为当环境中存在过量重金属时,细菌可通过渗透屏障、胞外蛋白/螯合剂与被动容忍作用减少对重金属离子的摄取,当重金属离子跨膜进入胞内后,细菌可通过外排系统将胞内重金属离子主动运输到胞外.此外,细菌还可通过重金属与胞内蛋白/螯合剂结合、酶解毒、降低重金属靶标敏感性等多重作用使重金属在胞内进行累积和解毒〔27,30〕. ...
... 〔28〕认为重金属抗性机制包括一般抗性机制和依赖金属的特殊抗性机制;J. A. LEMIRE等〔29〕把细菌的重金属抗性机制归纳为如图1所示的5个方面,包括细胞外隔离、减少摄取、外排系统、胞内累积和解毒机制,其认为当环境中存在过量重金属时,细菌可通过渗透屏障、胞外蛋白/螯合剂与被动容忍作用减少对重金属离子的摄取,当重金属离子跨膜进入胞内后,细菌可通过外排系统将胞内重金属离子主动运输到胞外.此外,细菌还可通过重金属与胞内蛋白/螯合剂结合、酶解毒、降低重金属靶标敏感性等多重作用使重金属在胞内进行累积和解毒〔27,30〕. ...
2
2004
... 为了避免过量重金属离子对细胞的生物毒性,细菌在长期的进化过程中形成了复杂多样的重金属抗性机制以维持正常的细胞内重金属离子浓度.依据不同的角度和理解,研究者对细菌的重金属抗性机制有着不同的划分〔27-28〕,如R. M. MAIER等〔28〕认为重金属抗性机制包括一般抗性机制和依赖金属的特殊抗性机制;J. A. LEMIRE等〔29〕把细菌的重金属抗性机制归纳为如图1所示的5个方面,包括细胞外隔离、减少摄取、外排系统、胞内累积和解毒机制,其认为当环境中存在过量重金属时,细菌可通过渗透屏障、胞外蛋白/螯合剂与被动容忍作用减少对重金属离子的摄取,当重金属离子跨膜进入胞内后,细菌可通过外排系统将胞内重金属离子主动运输到胞外.此外,细菌还可通过重金属与胞内蛋白/螯合剂结合、酶解毒、降低重金属靶标敏感性等多重作用使重金属在胞内进行累积和解毒〔27,30〕. ...
... 〔28〕认为重金属抗性机制包括一般抗性机制和依赖金属的特殊抗性机制;J. A. LEMIRE等〔29〕把细菌的重金属抗性机制归纳为如图1所示的5个方面,包括细胞外隔离、减少摄取、外排系统、胞内累积和解毒机制,其认为当环境中存在过量重金属时,细菌可通过渗透屏障、胞外蛋白/螯合剂与被动容忍作用减少对重金属离子的摄取,当重金属离子跨膜进入胞内后,细菌可通过外排系统将胞内重金属离子主动运输到胞外.此外,细菌还可通过重金属与胞内蛋白/螯合剂结合、酶解毒、降低重金属靶标敏感性等多重作用使重金属在胞内进行累积和解毒〔27,30〕. ...
Antimicrobial activity of metals:Mechanisms,molecular targets and applications
1
2013
... 为了避免过量重金属离子对细胞的生物毒性,细菌在长期的进化过程中形成了复杂多样的重金属抗性机制以维持正常的细胞内重金属离子浓度.依据不同的角度和理解,研究者对细菌的重金属抗性机制有着不同的划分〔27-28〕,如R. M. MAIER等〔28〕认为重金属抗性机制包括一般抗性机制和依赖金属的特殊抗性机制;J. A. LEMIRE等〔29〕把细菌的重金属抗性机制归纳为如图1所示的5个方面,包括细胞外隔离、减少摄取、外排系统、胞内累积和解毒机制,其认为当环境中存在过量重金属时,细菌可通过渗透屏障、胞外蛋白/螯合剂与被动容忍作用减少对重金属离子的摄取,当重金属离子跨膜进入胞内后,细菌可通过外排系统将胞内重金属离子主动运输到胞外.此外,细菌还可通过重金属与胞内蛋白/螯合剂结合、酶解毒、降低重金属靶标敏感性等多重作用使重金属在胞内进行累积和解毒〔27,30〕. ...
The confluence of heavy metal biooxidation and heavy metal resistance:Implications for bioleaching by extreme thermoacidophiles
1
2015
... 为了避免过量重金属离子对细胞的生物毒性,细菌在长期的进化过程中形成了复杂多样的重金属抗性机制以维持正常的细胞内重金属离子浓度.依据不同的角度和理解,研究者对细菌的重金属抗性机制有着不同的划分〔27-28〕,如R. M. MAIER等〔28〕认为重金属抗性机制包括一般抗性机制和依赖金属的特殊抗性机制;J. A. LEMIRE等〔29〕把细菌的重金属抗性机制归纳为如图1所示的5个方面,包括细胞外隔离、减少摄取、外排系统、胞内累积和解毒机制,其认为当环境中存在过量重金属时,细菌可通过渗透屏障、胞外蛋白/螯合剂与被动容忍作用减少对重金属离子的摄取,当重金属离子跨膜进入胞内后,细菌可通过外排系统将胞内重金属离子主动运输到胞外.此外,细菌还可通过重金属与胞内蛋白/螯合剂结合、酶解毒、降低重金属靶标敏感性等多重作用使重金属在胞内进行累积和解毒〔27,30〕. ...
微生物对重金属的抗性及解毒机理
1
1998
... 鉴于对于重金属胞内外排机制国内外已经有大量专门的综述类文章〔31-32〕,以下仅对细菌的重金属抗性机制中的生物吸附和胞内累积进行综述. ...
Microbial resistance to heavy metals and detoxification mechanism
1
1998
... 鉴于对于重金属胞内外排机制国内外已经有大量专门的综述类文章〔31-32〕,以下仅对细菌的重金属抗性机制中的生物吸附和胞内累积进行综述. ...
细菌对重金属的抗性及解毒机理研究进展
1
2015
... 鉴于对于重金属胞内外排机制国内外已经有大量专门的综述类文章〔31-32〕,以下仅对细菌的重金属抗性机制中的生物吸附和胞内累积进行综述. ...
On the research advances of the resistance and de-toxicity power of bacteria from heavy metals
1
2015
... 鉴于对于重金属胞内外排机制国内外已经有大量专门的综述类文章〔31-32〕,以下仅对细菌的重金属抗性机制中的生物吸附和胞内累积进行综述. ...
Highlighting inconsistencies regarding metal biosorption
1
2016
... 生物吸附(Biosorption)是一个概括性的术语,它包括多种作用机制,A. ROBALDS等〔33〕把生物吸附机理归纳为物理吸附、化学吸附、离子交换和微沉淀,其中化学吸附包括螯合作用与共价作用.对于重金属抗性菌而言,生物吸附是指其通过细胞外部和细胞表面的吸附位点将重金属离子吸附在细胞表面,减少细胞对重金属的摄入量,以提高自身抵抗重金属能力的重要机制〔34〕. ...
Role of extracellular polymeric substances in Cu(Ⅱ) adsorption on Bacillus subtilis and Pseudomonas putida
1
2011
... 生物吸附(Biosorption)是一个概括性的术语,它包括多种作用机制,A. ROBALDS等〔33〕把生物吸附机理归纳为物理吸附、化学吸附、离子交换和微沉淀,其中化学吸附包括螯合作用与共价作用.对于重金属抗性菌而言,生物吸附是指其通过细胞外部和细胞表面的吸附位点将重金属离子吸附在细胞表面,减少细胞对重金属的摄入量,以提高自身抵抗重金属能力的重要机制〔34〕. ...
Extracellular polymeric substances(EPS) of microbial aggregates in biological wastewater treatment systems:A review
1
2010
... 重金属细胞外部吸附的主体是胞外聚合物(Extracellular polymeric substance,EPS),它是微生物活细胞代谢过程分泌在细胞表层的高分子聚合物,主要成分为不定比例的蛋白质和多糖,以及少量的核酸、脂类和腐殖酸等〔35-36〕. ...
胞外聚合物及其对重金属吸附作用的研究进展
1
2019
... 重金属细胞外部吸附的主体是胞外聚合物(Extracellular polymeric substance,EPS),它是微生物活细胞代谢过程分泌在细胞表层的高分子聚合物,主要成分为不定比例的蛋白质和多糖,以及少量的核酸、脂类和腐殖酸等〔35-36〕. ...
Research progress in the extracellular polymeric substances and their adsorption effects on heavy metals
1
2019
... 重金属细胞外部吸附的主体是胞外聚合物(Extracellular polymeric substance,EPS),它是微生物活细胞代谢过程分泌在细胞表层的高分子聚合物,主要成分为不定比例的蛋白质和多糖,以及少量的核酸、脂类和腐殖酸等〔35-36〕. ...
微生物抗重金属的生理机制
1
2009
... EPS中的大分子含有多种可以与重金属离子结合的阴离子基团,如羧基、羟基、巯基和磷酸根基团等,各阴离子基团通过配位、吸附、沉淀和架桥絮凝等作用吸附重金属离子,防止重金属离子与细胞作用〔37〕.研究发现适当的重金属离子浓度可以诱导细菌增加EPS的分泌,即在一定浓度的重金属环境中,细菌对重金属胁迫产生响应而上调胞外聚合物基因的表达,从而增加胞外聚合物对重金属离子的捕获,如Pseudomonas putida〔26〕、Pseudomonas aeruginosa〔38〕、B. subtilis〔39〕在铜的作用下,EPS都有明显的增加.反之,如果去除菌属细菌产生的胞外多糖,细菌吸附重金属能力明显降低,对重金属的敏感性增加. ...
The physiological mechanism of microbial heavy metals tolerance
1
2009
... EPS中的大分子含有多种可以与重金属离子结合的阴离子基团,如羧基、羟基、巯基和磷酸根基团等,各阴离子基团通过配位、吸附、沉淀和架桥絮凝等作用吸附重金属离子,防止重金属离子与细胞作用〔37〕.研究发现适当的重金属离子浓度可以诱导细菌增加EPS的分泌,即在一定浓度的重金属环境中,细菌对重金属胁迫产生响应而上调胞外聚合物基因的表达,从而增加胞外聚合物对重金属离子的捕获,如Pseudomonas putida〔26〕、Pseudomonas aeruginosa〔38〕、B. subtilis〔39〕在铜的作用下,EPS都有明显的增加.反之,如果去除菌属细菌产生的胞外多糖,细菌吸附重金属能力明显降低,对重金属的敏感性增加. ...
Heavy metal resistance of biofilm and planktonic Pseudomonas aeruginosa
1
2003
... EPS中的大分子含有多种可以与重金属离子结合的阴离子基团,如羧基、羟基、巯基和磷酸根基团等,各阴离子基团通过配位、吸附、沉淀和架桥絮凝等作用吸附重金属离子,防止重金属离子与细胞作用〔37〕.研究发现适当的重金属离子浓度可以诱导细菌增加EPS的分泌,即在一定浓度的重金属环境中,细菌对重金属胁迫产生响应而上调胞外聚合物基因的表达,从而增加胞外聚合物对重金属离子的捕获,如Pseudomonas putida〔26〕、Pseudomonas aeruginosa〔38〕、B. subtilis〔39〕在铜的作用下,EPS都有明显的增加.反之,如果去除菌属细菌产生的胞外多糖,细菌吸附重金属能力明显降低,对重金属的敏感性增加. ...
Biosorption mechanisms of Cu(Ⅱ) by extracellular polymeric substancesfrom Bacillus subtilis
1
2014
... EPS中的大分子含有多种可以与重金属离子结合的阴离子基团,如羧基、羟基、巯基和磷酸根基团等,各阴离子基团通过配位、吸附、沉淀和架桥絮凝等作用吸附重金属离子,防止重金属离子与细胞作用〔37〕.研究发现适当的重金属离子浓度可以诱导细菌增加EPS的分泌,即在一定浓度的重金属环境中,细菌对重金属胁迫产生响应而上调胞外聚合物基因的表达,从而增加胞外聚合物对重金属离子的捕获,如Pseudomonas putida〔26〕、Pseudomonas aeruginosa〔38〕、B. subtilis〔39〕在铜的作用下,EPS都有明显的增加.反之,如果去除菌属细菌产生的胞外多糖,细菌吸附重金属能力明显降低,对重金属的敏感性增加. ...
A comparative review towards potential of microbial cells for heavy metal removal with emphasis on biosorption and bioaccumulation
1
2016
... 随着研究的不断深入,细胞外部吸附和细胞表面吸附机理逐渐明晰.研究表明,除了EPS,构成细菌细胞壁的化学组分也可以吸附重金属离子.这是因为细菌细胞壁组分中所含的大量肽聚糖、脂多糖、脂类和磷壁酸等也可与重金属离子发生物理、化学作用,通过离子交换、表面络合、氧化还原、无机微沉淀和静电作用等吸附重金属离子从而减少进入胞内的重金属离子.如革兰氏阳性菌的细胞壁中含有90%的肽聚糖以及较多的磷壁酸,带有较强的负电荷,能吸附重金属阳离子,而革兰氏阴性菌细胞壁中脂多糖内的磷酸基团和磷脂已被证明是与重金属相互作用的主要活性位点〔40〕.需要指出的是,细胞表面吸附是不依赖微生物代谢活动的被动吸附过程,即使是微生物的死细胞,其生物质作为吸附剂同样可以吸附重金属离子. ...
乳酸菌对重金属污染的生物修复作用
1
2014
... 细菌对重金属的生物累积存在2种可能机制.一种是重金属离子通过细胞膜进入到细胞质中,在细胞内特定的区域被隔离,以防止其接触到重要的细胞组分或细胞器〔41〕,另一种是重金属离子与细菌体内有生物活性的配位体发生特异的生物化学反应后形成螯合物对细菌进行生物解毒〔42〕.原核生物(如细菌)胞内含有谷胱甘肽(Glutathione,GSH)、金属硫蛋白等,这些物质可与重金属离子结合形成螯合物,对于重金属的解毒和防御重金属引起的氧化胁迫具有重要作用.GSH是由谷氨酸、半胱氨酸及甘氨酸组成的三肽,其含有的氨基、巯基、羧基和酰胺基等多种配位基团可与重金属离子结合,在生物体内对重金属进行累积.GSH还可以清除因重金属胁迫产生的活性氧自由基,以此来降低重金属离子对细胞的生物毒性〔43〕.细菌的重金属抗性机制是菌细胞应对重金属胁迫产生的一系列复杂的生理生化反应,Ning HUANG等〔5〕通过基因组与转录组学研究表明细菌对重金属的抗性往往不依赖于单一的机制,而是多种系统相互作用的结果. ...
Bioremediation of heavy metal contamination by Lactic acid bacteria
1
2014
... 细菌对重金属的生物累积存在2种可能机制.一种是重金属离子通过细胞膜进入到细胞质中,在细胞内特定的区域被隔离,以防止其接触到重要的细胞组分或细胞器〔41〕,另一种是重金属离子与细菌体内有生物活性的配位体发生特异的生物化学反应后形成螯合物对细菌进行生物解毒〔42〕.原核生物(如细菌)胞内含有谷胱甘肽(Glutathione,GSH)、金属硫蛋白等,这些物质可与重金属离子结合形成螯合物,对于重金属的解毒和防御重金属引起的氧化胁迫具有重要作用.GSH是由谷氨酸、半胱氨酸及甘氨酸组成的三肽,其含有的氨基、巯基、羧基和酰胺基等多种配位基团可与重金属离子结合,在生物体内对重金属进行累积.GSH还可以清除因重金属胁迫产生的活性氧自由基,以此来降低重金属离子对细胞的生物毒性〔43〕.细菌的重金属抗性机制是菌细胞应对重金属胁迫产生的一系列复杂的生理生化反应,Ning HUANG等〔5〕通过基因组与转录组学研究表明细菌对重金属的抗性往往不依赖于单一的机制,而是多种系统相互作用的结果. ...
Effects of copper ions on the free radical-scavenging properties of reduced gluthathione:Implications of a complex formation
1
2000
... 细菌对重金属的生物累积存在2种可能机制.一种是重金属离子通过细胞膜进入到细胞质中,在细胞内特定的区域被隔离,以防止其接触到重要的细胞组分或细胞器〔41〕,另一种是重金属离子与细菌体内有生物活性的配位体发生特异的生物化学反应后形成螯合物对细菌进行生物解毒〔42〕.原核生物(如细菌)胞内含有谷胱甘肽(Glutathione,GSH)、金属硫蛋白等,这些物质可与重金属离子结合形成螯合物,对于重金属的解毒和防御重金属引起的氧化胁迫具有重要作用.GSH是由谷氨酸、半胱氨酸及甘氨酸组成的三肽,其含有的氨基、巯基、羧基和酰胺基等多种配位基团可与重金属离子结合,在生物体内对重金属进行累积.GSH还可以清除因重金属胁迫产生的活性氧自由基,以此来降低重金属离子对细胞的生物毒性〔43〕.细菌的重金属抗性机制是菌细胞应对重金属胁迫产生的一系列复杂的生理生化反应,Ning HUANG等〔5〕通过基因组与转录组学研究表明细菌对重金属的抗性往往不依赖于单一的机制,而是多种系统相互作用的结果. ...
微生物与重金属的相互作用机理研究进展
1
2007
... 细菌对重金属的生物累积存在2种可能机制.一种是重金属离子通过细胞膜进入到细胞质中,在细胞内特定的区域被隔离,以防止其接触到重要的细胞组分或细胞器〔41〕,另一种是重金属离子与细菌体内有生物活性的配位体发生特异的生物化学反应后形成螯合物对细菌进行生物解毒〔42〕.原核生物(如细菌)胞内含有谷胱甘肽(Glutathione,GSH)、金属硫蛋白等,这些物质可与重金属离子结合形成螯合物,对于重金属的解毒和防御重金属引起的氧化胁迫具有重要作用.GSH是由谷氨酸、半胱氨酸及甘氨酸组成的三肽,其含有的氨基、巯基、羧基和酰胺基等多种配位基团可与重金属离子结合,在生物体内对重金属进行累积.GSH还可以清除因重金属胁迫产生的活性氧自由基,以此来降低重金属离子对细胞的生物毒性〔43〕.细菌的重金属抗性机制是菌细胞应对重金属胁迫产生的一系列复杂的生理生化反应,Ning HUANG等〔5〕通过基因组与转录组学研究表明细菌对重金属的抗性往往不依赖于单一的机制,而是多种系统相互作用的结果. ...
Studies on the mechanisms of interaction between microboes and heavy metals
1
2007
... 细菌对重金属的生物累积存在2种可能机制.一种是重金属离子通过细胞膜进入到细胞质中,在细胞内特定的区域被隔离,以防止其接触到重要的细胞组分或细胞器〔41〕,另一种是重金属离子与细菌体内有生物活性的配位体发生特异的生物化学反应后形成螯合物对细菌进行生物解毒〔42〕.原核生物(如细菌)胞内含有谷胱甘肽(Glutathione,GSH)、金属硫蛋白等,这些物质可与重金属离子结合形成螯合物,对于重金属的解毒和防御重金属引起的氧化胁迫具有重要作用.GSH是由谷氨酸、半胱氨酸及甘氨酸组成的三肽,其含有的氨基、巯基、羧基和酰胺基等多种配位基团可与重金属离子结合,在生物体内对重金属进行累积.GSH还可以清除因重金属胁迫产生的活性氧自由基,以此来降低重金属离子对细胞的生物毒性〔43〕.细菌的重金属抗性机制是菌细胞应对重金属胁迫产生的一系列复杂的生理生化反应,Ning HUANG等〔5〕通过基因组与转录组学研究表明细菌对重金属的抗性往往不依赖于单一的机制,而是多种系统相互作用的结果. ...
Preparation of carbon fibers/carbon/alumina tubular composite membranes and their applications in treating Cu-CMP wastewater by a novel electrochemical process
1
2008
... 铜是应用最广泛的重金属之一,含铜废水是重金属废水的典型代表.传统的含铜废水主要来自铜矿采选、冶炼、化工、印染、电镀等行业排放.近年来,随着新兴电子产业的发展,印刷电路板、集成电路制造行业生产过程中产生大量低浓度含铜废水〔ρ(Cu2+)<100 mg/L〕,此类废水的处理日益受到研究者的关注.化学机械研磨(CMP)已成为半导体集成电路制造过程中实现晶圆全局均匀平坦化的关键工艺,该工艺需要大量超纯水清洗晶圆研磨过程中产生的铜离子、二氧化硅及多种化学药剂.因此,CMP废水排放量大,约占整个半导体制造过程废水总量的30%~40%〔44〕,且其水质成分复杂,悬浮物和Cu2+含量较高〔45-48〕. ...
Synergistic effects of citric acid and polyethyleneimine to remove copper from aqueous solutions
1
2009
... 铜是应用最广泛的重金属之一,含铜废水是重金属废水的典型代表.传统的含铜废水主要来自铜矿采选、冶炼、化工、印染、电镀等行业排放.近年来,随着新兴电子产业的发展,印刷电路板、集成电路制造行业生产过程中产生大量低浓度含铜废水〔ρ(Cu2+)<100 mg/L〕,此类废水的处理日益受到研究者的关注.化学机械研磨(CMP)已成为半导体集成电路制造过程中实现晶圆全局均匀平坦化的关键工艺,该工艺需要大量超纯水清洗晶圆研磨过程中产生的铜离子、二氧化硅及多种化学药剂.因此,CMP废水排放量大,约占整个半导体制造过程废水总量的30%~40%〔44〕,且其水质成分复杂,悬浮物和Cu2+含量较高〔45-48〕. ...
Performance of integrated membrane filtration and electrodialysis processes for copper recovery from wafer polishing wastewater
0
2014
Study of COD and turbidity removal from real oxide-CMP wastewater by iron electrocoagulation and the evaluation of specific energy consumption
0
2009
Bioremoval of Cu2+ from CMP wastewater by a novel copper-resistant bacterium Cupriavidus gilardii CR3:Characteristics and mechanisms
1
2017
... 铜是应用最广泛的重金属之一,含铜废水是重金属废水的典型代表.传统的含铜废水主要来自铜矿采选、冶炼、化工、印染、电镀等行业排放.近年来,随着新兴电子产业的发展,印刷电路板、集成电路制造行业生产过程中产生大量低浓度含铜废水〔ρ(Cu2+)<100 mg/L〕,此类废水的处理日益受到研究者的关注.化学机械研磨(CMP)已成为半导体集成电路制造过程中实现晶圆全局均匀平坦化的关键工艺,该工艺需要大量超纯水清洗晶圆研磨过程中产生的铜离子、二氧化硅及多种化学药剂.因此,CMP废水排放量大,约占整个半导体制造过程废水总量的30%~40%〔44〕,且其水质成分复杂,悬浮物和Cu2+含量较高〔45-48〕. ...
化学机械研磨废水处理及回用技术的研究进展
1
2012
... 目前,CMP废水中Cu2+的处理方法主要有化学沉淀法、电化学方法〔49〕、离子交换树脂法〔50〕和膜过滤处理法〔51〕.这些物理化学处理方法可实现Cu2+达标排放,但缺点是耗能大、费用高.因此,国内外研究者尝试采用生物法处理CMP废水中的Cu2+〔52-57〕. ...
Progress on treatment and reuse technologies of chemical mechanical polishing(CMP) wastewater
1
2012
... 目前,CMP废水中Cu2+的处理方法主要有化学沉淀法、电化学方法〔49〕、离子交换树脂法〔50〕和膜过滤处理法〔51〕.这些物理化学处理方法可实现Cu2+达标排放,但缺点是耗能大、费用高.因此,国内外研究者尝试采用生物法处理CMP废水中的Cu2+〔52-57〕. ...
Treatment of chemical mechanical polishing wastewater by electrocoagulation:System performances and sludge settling characteristics
1
2004
... 目前,CMP废水中Cu2+的处理方法主要有化学沉淀法、电化学方法〔49〕、离子交换树脂法〔50〕和膜过滤处理法〔51〕.这些物理化学处理方法可实现Cu2+达标排放,但缺点是耗能大、费用高.因此,国内外研究者尝试采用生物法处理CMP废水中的Cu2+〔52-57〕. ...
1
... 目前,CMP废水中Cu2+的处理方法主要有化学沉淀法、电化学方法〔49〕、离子交换树脂法〔50〕和膜过滤处理法〔51〕.这些物理化学处理方法可实现Cu2+达标排放,但缺点是耗能大、费用高.因此,国内外研究者尝试采用生物法处理CMP废水中的Cu2+〔52-57〕. ...
1
... 目前,CMP废水中Cu2+的处理方法主要有化学沉淀法、电化学方法〔49〕、离子交换树脂法〔50〕和膜过滤处理法〔51〕.这些物理化学处理方法可实现Cu2+达标排放,但缺点是耗能大、费用高.因此,国内外研究者尝试采用生物法处理CMP废水中的Cu2+〔52-57〕. ...
Biosorption of copper(Ⅱ) from chemical mechanical planarization wastewaters
2
2003
... 在去除CMP废水中Cu2+的研究中,所采用的重金属抗性菌有固定化〔52-56〕和浮游态〔57〕2种形式.从已有研究来看,绝大多数研究对细菌采用了固定化方式.一般来说,固定化微生物技术可使微生物固着生长,保持高的生物活性,是提高重金属抗性微生物活性的重要方法.国内外有大量关于采用生物膜及其胞外聚合物对Cu2+进行吸附处理的报道〔58〕. ...
... L. C. STANLEY等〔52〕以分离自美国亚利桑那州绿谷市Cyprus Sierrita铜矿周边土壤的细菌Staphylococcus sp. Soil 5Y为重金属抗性菌,在进水流量为0.8~1.6 mL/min条件下,采用微生物填充柱连续吸附去除模拟和真实CMP废水中的Cu2+〔ρ(Cu2+)=10~40 mg/L〕,实验结果表明,该方法可以有效去除CMP废水中Cu2+,使其出水质量浓度低于1 mg/L,细菌Staphylococcus sp. Soil 5Y对于Cu2+的吸附量为(140±50) mg/g,且其再生过程无需处理大量的有害固废.A. P. MOSIER等〔53〕和Xiaoyu WANG等〔54〕同样采用类似的重金属抗性菌活细胞固定微生物吸附柱研究了重金属抗性菌对于CMP废水中Cu2+的去除.A. P. MOSIER等〔53〕所采用的抗性微生物是重金属抗性细菌Lactobacillus casei和毕赤酵母Pichia pastoris的混合菌生物膜,细菌的固定化载体为活性炭,实验表明固定化混合生物膜对含5 mg/L Cu2+的模拟CMP废水中Cu2+的去除率为79%.此外,该研究采用1.0 mmol/L的HCl溶液对吸附Cu2+饱和后的微生物吸附柱进行冲洗,考察生物膜的再生功能,结果表明固定化生物膜具有再生和重复使用的潜力.Xiaoyu WANG等〔54〕采用的模拟CMP废水成分除了Cu2+之外,还考虑了CMP工艺过程中存在的大量纳米二氧化硅.实验中所采用的重金属抗性菌为C. gilardii CR3,菌株CR3的固定化载体为石英砂.实验结果表明,当CMP废水中伴有纳米二氧化硅时不影响细菌对Cu2+的吸附,相较于Langmuir模型,Freundlich模型对于该吸附行为的拟合度更高(R2 = 0.99),但是当CMP废水中无纳米二氧化硅时,Langmuir(R2 =0.99)模型则更适合细菌对于Cu2+吸附行为的描述. ...
Microbial biofilms for the removal of Cu2+ from CMP wastewater
2
2015
... L. C. STANLEY等〔52〕以分离自美国亚利桑那州绿谷市Cyprus Sierrita铜矿周边土壤的细菌Staphylococcus sp. Soil 5Y为重金属抗性菌,在进水流量为0.8~1.6 mL/min条件下,采用微生物填充柱连续吸附去除模拟和真实CMP废水中的Cu2+〔ρ(Cu2+)=10~40 mg/L〕,实验结果表明,该方法可以有效去除CMP废水中Cu2+,使其出水质量浓度低于1 mg/L,细菌Staphylococcus sp. Soil 5Y对于Cu2+的吸附量为(140±50) mg/g,且其再生过程无需处理大量的有害固废.A. P. MOSIER等〔53〕和Xiaoyu WANG等〔54〕同样采用类似的重金属抗性菌活细胞固定微生物吸附柱研究了重金属抗性菌对于CMP废水中Cu2+的去除.A. P. MOSIER等〔53〕所采用的抗性微生物是重金属抗性细菌Lactobacillus casei和毕赤酵母Pichia pastoris的混合菌生物膜,细菌的固定化载体为活性炭,实验表明固定化混合生物膜对含5 mg/L Cu2+的模拟CMP废水中Cu2+的去除率为79%.此外,该研究采用1.0 mmol/L的HCl溶液对吸附Cu2+饱和后的微生物吸附柱进行冲洗,考察生物膜的再生功能,结果表明固定化生物膜具有再生和重复使用的潜力.Xiaoyu WANG等〔54〕采用的模拟CMP废水成分除了Cu2+之外,还考虑了CMP工艺过程中存在的大量纳米二氧化硅.实验中所采用的重金属抗性菌为C. gilardii CR3,菌株CR3的固定化载体为石英砂.实验结果表明,当CMP废水中伴有纳米二氧化硅时不影响细菌对Cu2+的吸附,相较于Langmuir模型,Freundlich模型对于该吸附行为的拟合度更高(R2 = 0.99),但是当CMP废水中无纳米二氧化硅时,Langmuir(R2 =0.99)模型则更适合细菌对于Cu2+吸附行为的描述. ...
... 〔53〕所采用的抗性微生物是重金属抗性细菌Lactobacillus casei和毕赤酵母Pichia pastoris的混合菌生物膜,细菌的固定化载体为活性炭,实验表明固定化混合生物膜对含5 mg/L Cu2+的模拟CMP废水中Cu2+的去除率为79%.此外,该研究采用1.0 mmol/L的HCl溶液对吸附Cu2+饱和后的微生物吸附柱进行冲洗,考察生物膜的再生功能,结果表明固定化生物膜具有再生和重复使用的潜力.Xiaoyu WANG等〔54〕采用的模拟CMP废水成分除了Cu2+之外,还考虑了CMP工艺过程中存在的大量纳米二氧化硅.实验中所采用的重金属抗性菌为C. gilardii CR3,菌株CR3的固定化载体为石英砂.实验结果表明,当CMP废水中伴有纳米二氧化硅时不影响细菌对Cu2+的吸附,相较于Langmuir模型,Freundlich模型对于该吸附行为的拟合度更高(R2 = 0.99),但是当CMP废水中无纳米二氧化硅时,Langmuir(R2 =0.99)模型则更适合细菌对于Cu2+吸附行为的描述. ...
Copper removal from semiconductor CMP wastewater in the presence of nano-SiO2 through biosorption
2
2021
... L. C. STANLEY等〔52〕以分离自美国亚利桑那州绿谷市Cyprus Sierrita铜矿周边土壤的细菌Staphylococcus sp. Soil 5Y为重金属抗性菌,在进水流量为0.8~1.6 mL/min条件下,采用微生物填充柱连续吸附去除模拟和真实CMP废水中的Cu2+〔ρ(Cu2+)=10~40 mg/L〕,实验结果表明,该方法可以有效去除CMP废水中Cu2+,使其出水质量浓度低于1 mg/L,细菌Staphylococcus sp. Soil 5Y对于Cu2+的吸附量为(140±50) mg/g,且其再生过程无需处理大量的有害固废.A. P. MOSIER等〔53〕和Xiaoyu WANG等〔54〕同样采用类似的重金属抗性菌活细胞固定微生物吸附柱研究了重金属抗性菌对于CMP废水中Cu2+的去除.A. P. MOSIER等〔53〕所采用的抗性微生物是重金属抗性细菌Lactobacillus casei和毕赤酵母Pichia pastoris的混合菌生物膜,细菌的固定化载体为活性炭,实验表明固定化混合生物膜对含5 mg/L Cu2+的模拟CMP废水中Cu2+的去除率为79%.此外,该研究采用1.0 mmol/L的HCl溶液对吸附Cu2+饱和后的微生物吸附柱进行冲洗,考察生物膜的再生功能,结果表明固定化生物膜具有再生和重复使用的潜力.Xiaoyu WANG等〔54〕采用的模拟CMP废水成分除了Cu2+之外,还考虑了CMP工艺过程中存在的大量纳米二氧化硅.实验中所采用的重金属抗性菌为C. gilardii CR3,菌株CR3的固定化载体为石英砂.实验结果表明,当CMP废水中伴有纳米二氧化硅时不影响细菌对Cu2+的吸附,相较于Langmuir模型,Freundlich模型对于该吸附行为的拟合度更高(R2 = 0.99),但是当CMP废水中无纳米二氧化硅时,Langmuir(R2 =0.99)模型则更适合细菌对于Cu2+吸附行为的描述. ...
... 〔54〕采用的模拟CMP废水成分除了Cu2+之外,还考虑了CMP工艺过程中存在的大量纳米二氧化硅.实验中所采用的重金属抗性菌为C. gilardii CR3,菌株CR3的固定化载体为石英砂.实验结果表明,当CMP废水中伴有纳米二氧化硅时不影响细菌对Cu2+的吸附,相较于Langmuir模型,Freundlich模型对于该吸附行为的拟合度更高(R2 = 0.99),但是当CMP废水中无纳米二氧化硅时,Langmuir(R2 =0.99)模型则更适合细菌对于Cu2+吸附行为的描述. ...
Bacterial biofilms for sequestration of Cu2+ from CMP wastewater
0
2014
Biotreatment of copper and isopropyl alcohol in waste from semiconductor manufacturing
1
2004
... 在去除CMP废水中Cu2+的研究中,所采用的重金属抗性菌有固定化〔52-56〕和浮游态〔57〕2种形式.从已有研究来看,绝大多数研究对细菌采用了固定化方式.一般来说,固定化微生物技术可使微生物固着生长,保持高的生物活性,是提高重金属抗性微生物活性的重要方法.国内外有大量关于采用生物膜及其胞外聚合物对Cu2+进行吸附处理的报道〔58〕. ...
Bioremoval of Cu2+ from CMP wastewater by a novel copper-resistant bacterium Cupriavidus gilardii CR3:Characteristics and mechanisms
1
2017
... 在去除CMP废水中Cu2+的研究中,所采用的重金属抗性菌有固定化〔52-56〕和浮游态〔57〕2种形式.从已有研究来看,绝大多数研究对细菌采用了固定化方式.一般来说,固定化微生物技术可使微生物固着生长,保持高的生物活性,是提高重金属抗性微生物活性的重要方法.国内外有大量关于采用生物膜及其胞外聚合物对Cu2+进行吸附处理的报道〔58〕. ...
Cu2+对生物膜及其胞外聚合物的影响
1
2014
... 在去除CMP废水中Cu2+的研究中,所采用的重金属抗性菌有固定化〔52-56〕和浮游态〔57〕2种形式.从已有研究来看,绝大多数研究对细菌采用了固定化方式.一般来说,固定化微生物技术可使微生物固着生长,保持高的生物活性,是提高重金属抗性微生物活性的重要方法.国内外有大量关于采用生物膜及其胞外聚合物对Cu2+进行吸附处理的报道〔58〕. ...
Effect of Cu2+ on biofilm and extracellular polymeric substance
1
2014
... 在去除CMP废水中Cu2+的研究中,所采用的重金属抗性菌有固定化〔52-56〕和浮游态〔57〕2种形式.从已有研究来看,绝大多数研究对细菌采用了固定化方式.一般来说,固定化微生物技术可使微生物固着生长,保持高的生物活性,是提高重金属抗性微生物活性的重要方法.国内外有大量关于采用生物膜及其胞外聚合物对Cu2+进行吸附处理的报道〔58〕. ...






 津公网安备 12010602120337号
津公网安备 12010602120337号{kind=link}
{kind=link}
{kind=link}
{kind=link}