丁辛醇生产废水处理技术研究
1
2007
... 煤化工的DAVY/DOW低压羰基合成工艺生产丁辛醇时会产生大量废水〔1〕.丁辛醇废水(BOW)含有的有机物复杂、浓度高,且具有强碱性、高盐度,属于典型的难降解有机工业废水.目前BOW的主流处理工艺以萃取法、焚烧法、空气催化氧化法等物化法为主,但这些处理方法的缺点在于处理流程复杂、成本高〔2〕.因此亟需一种绿色、高效的BOW处理技术. ...
丁辛醇生产废水处理技术研究
1
2007
... 煤化工的DAVY/DOW低压羰基合成工艺生产丁辛醇时会产生大量废水〔1〕.丁辛醇废水(BOW)含有的有机物复杂、浓度高,且具有强碱性、高盐度,属于典型的难降解有机工业废水.目前BOW的主流处理工艺以萃取法、焚烧法、空气催化氧化法等物化法为主,但这些处理方法的缺点在于处理流程复杂、成本高〔2〕.因此亟需一种绿色、高效的BOW处理技术. ...
丁辛醇缩合废水处理工艺研究
1
2020
... 煤化工的DAVY/DOW低压羰基合成工艺生产丁辛醇时会产生大量废水〔1〕.丁辛醇废水(BOW)含有的有机物复杂、浓度高,且具有强碱性、高盐度,属于典型的难降解有机工业废水.目前BOW的主流处理工艺以萃取法、焚烧法、空气催化氧化法等物化法为主,但这些处理方法的缺点在于处理流程复杂、成本高〔2〕.因此亟需一种绿色、高效的BOW处理技术. ...
丁辛醇缩合废水处理工艺研究
1
2020
... 煤化工的DAVY/DOW低压羰基合成工艺生产丁辛醇时会产生大量废水〔1〕.丁辛醇废水(BOW)含有的有机物复杂、浓度高,且具有强碱性、高盐度,属于典型的难降解有机工业废水.目前BOW的主流处理工艺以萃取法、焚烧法、空气催化氧化法等物化法为主,但这些处理方法的缺点在于处理流程复杂、成本高〔2〕.因此亟需一种绿色、高效的BOW处理技术. ...
Anaerobic digestion in global bio-energy production:Potential and research challenges
1
2011
... 厌氧消化是实现污染物降解和资源化的最有效途径之一〔3〕.通过厌氧消化BOW产生甲烷是较好的选择.然而BOW中高浓度的有机物和循环回用装置中积累的高盐分都会严重影响厌氧微生物的代谢能力,对厌氧系统的稳定性造成严重威胁.有研究提出将颗粒活性炭(GAC)应用于BOW厌氧消化,有利于解决甲烷产率低的问题〔4〕.由于GAC的高导电性能促进微生物间的直接种间电子转移(DIET)〔5〕,与使用传统电子载体(氢、甲酸等)进行物种间电子转移相比,DIET提高了甲烷生成效率〔6〕.GAC已被广泛应用于厌氧反应器中处理有毒难降解有机废水,且大多研究集中在UASB类型的反应器.相比之下,具备多级污泥床形式的厌氧折板反应器(ABR)具有更强的稳定性,更有利于GAC对电活性互营微生物的富集.然而,关于GAC强化ABR运行的研究较为少见. ...
Promoting direct inter species electron transfer and acetoclastic methanogenesis for enhancing anaerobic digestion of butanol octanol wastewater by coupling granular activated carbon and exogenous hydrogen
1
2021
... 厌氧消化是实现污染物降解和资源化的最有效途径之一〔3〕.通过厌氧消化BOW产生甲烷是较好的选择.然而BOW中高浓度的有机物和循环回用装置中积累的高盐分都会严重影响厌氧微生物的代谢能力,对厌氧系统的稳定性造成严重威胁.有研究提出将颗粒活性炭(GAC)应用于BOW厌氧消化,有利于解决甲烷产率低的问题〔4〕.由于GAC的高导电性能促进微生物间的直接种间电子转移(DIET)〔5〕,与使用传统电子载体(氢、甲酸等)进行物种间电子转移相比,DIET提高了甲烷生成效率〔6〕.GAC已被广泛应用于厌氧反应器中处理有毒难降解有机废水,且大多研究集中在UASB类型的反应器.相比之下,具备多级污泥床形式的厌氧折板反应器(ABR)具有更强的稳定性,更有利于GAC对电活性互营微生物的富集.然而,关于GAC强化ABR运行的研究较为少见. ...
Conductive material engineered direct inter species electron transfer(DIET) in anaerobic digestion:Mechanism and application
1
2020
... 厌氧消化是实现污染物降解和资源化的最有效途径之一〔3〕.通过厌氧消化BOW产生甲烷是较好的选择.然而BOW中高浓度的有机物和循环回用装置中积累的高盐分都会严重影响厌氧微生物的代谢能力,对厌氧系统的稳定性造成严重威胁.有研究提出将颗粒活性炭(GAC)应用于BOW厌氧消化,有利于解决甲烷产率低的问题〔4〕.由于GAC的高导电性能促进微生物间的直接种间电子转移(DIET)〔5〕,与使用传统电子载体(氢、甲酸等)进行物种间电子转移相比,DIET提高了甲烷生成效率〔6〕.GAC已被广泛应用于厌氧反应器中处理有毒难降解有机废水,且大多研究集中在UASB类型的反应器.相比之下,具备多级污泥床形式的厌氧折板反应器(ABR)具有更强的稳定性,更有利于GAC对电活性互营微生物的富集.然而,关于GAC强化ABR运行的研究较为少见. ...
Magnetite particles triggering a faster and more robust syntrophic pathway of methanogenic propionate degradation
1
2014
... 厌氧消化是实现污染物降解和资源化的最有效途径之一〔3〕.通过厌氧消化BOW产生甲烷是较好的选择.然而BOW中高浓度的有机物和循环回用装置中积累的高盐分都会严重影响厌氧微生物的代谢能力,对厌氧系统的稳定性造成严重威胁.有研究提出将颗粒活性炭(GAC)应用于BOW厌氧消化,有利于解决甲烷产率低的问题〔4〕.由于GAC的高导电性能促进微生物间的直接种间电子转移(DIET)〔5〕,与使用传统电子载体(氢、甲酸等)进行物种间电子转移相比,DIET提高了甲烷生成效率〔6〕.GAC已被广泛应用于厌氧反应器中处理有毒难降解有机废水,且大多研究集中在UASB类型的反应器.相比之下,具备多级污泥床形式的厌氧折板反应器(ABR)具有更强的稳定性,更有利于GAC对电活性互营微生物的富集.然而,关于GAC强化ABR运行的研究较为少见. ...
Treatment of organic pollution in industrial saline wastewater:A literature review
1
2006
... 在工业废水厌氧处理中,盐度也是影响厌氧微生物生长的一个主要因素〔7〕.高盐环境中的高渗透压会导致微生物细胞死亡〔8〕.Qian CHEN等〔9〕通过添加磁铁矿来富集假单胞菌,以增强高盐有机废水的厌氧消化,但DIET微生物耐盐性的研究仍然较少.盐度对于互营微生物富集的潜在影响值得探讨.笔者探究了高盐条件下厌氧颗粒活性炭折板工艺(GAC-ABR)处理BOW的效能变化,阐述了GAC-ABR对污泥特性的影响并对微生物群落结构进行分析. ...
Microbial community redundancy in anaerobic digestion drives process recovery after salinity exposure
1
2017
... 在工业废水厌氧处理中,盐度也是影响厌氧微生物生长的一个主要因素〔7〕.高盐环境中的高渗透压会导致微生物细胞死亡〔8〕.Qian CHEN等〔9〕通过添加磁铁矿来富集假单胞菌,以增强高盐有机废水的厌氧消化,但DIET微生物耐盐性的研究仍然较少.盐度对于互营微生物富集的潜在影响值得探讨.笔者探究了高盐条件下厌氧颗粒活性炭折板工艺(GAC-ABR)处理BOW的效能变化,阐述了GAC-ABR对污泥特性的影响并对微生物群落结构进行分析. ...
Magnetite enhances anaerobic digestion of high salinity organic wastewater
1
2020
... 在工业废水厌氧处理中,盐度也是影响厌氧微生物生长的一个主要因素〔7〕.高盐环境中的高渗透压会导致微生物细胞死亡〔8〕.Qian CHEN等〔9〕通过添加磁铁矿来富集假单胞菌,以增强高盐有机废水的厌氧消化,但DIET微生物耐盐性的研究仍然较少.盐度对于互营微生物富集的潜在影响值得探讨.笔者探究了高盐条件下厌氧颗粒活性炭折板工艺(GAC-ABR)处理BOW的效能变化,阐述了GAC-ABR对污泥特性的影响并对微生物群落结构进行分析. ...
Performance robustness of the UASB reactors treating saline phenolic wastewater and analysis of microbial community structure
2
2017
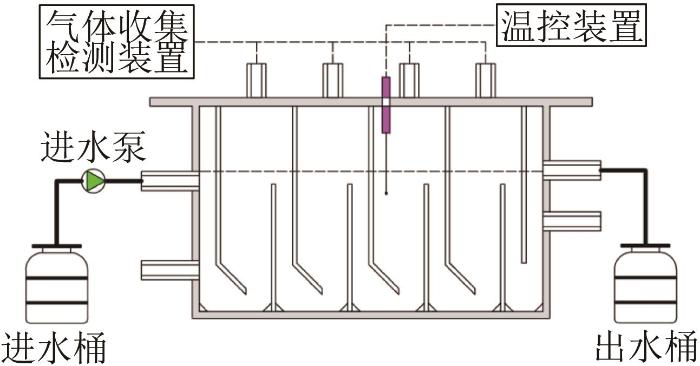
... 接种的活性污泥取自合肥某污水处理厂的SBR池,在4 ℃下储存.污泥的总悬浮物固体(TSS)为20.7 g/L,VSS/SS为0.55.GAC粒径为1 mm(18目,国药化学试剂有限公司).BOW取自曙光化学(集团)有限公司,主要组成见表1.添加的营养成分与前期研究〔10〕相同,所用试剂均为分析纯. ...
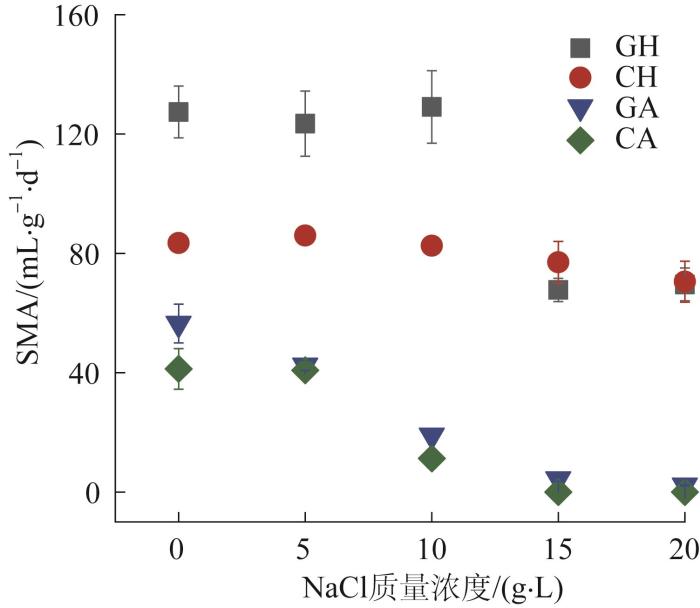
... 以乙酸钠(按COD计,2 000 mg/L)和H2/CO2(140 mL/35 mL)为底物,分别测定污泥的嗜乙酸和嗜氢产甲烷活性.污泥质量与底物COD的比值为1∶1,营养元素组成参考文献〔10〕.用高纯度氮气(99.99%)吹洗血清瓶约1~2 min,立即密封,在35 ℃、150 r/min的摇床中培养,根据甲烷产量随时间变化情况计算比产甲烷活性(SMA).按照上述方法测定0、5、10、15、20 g/L NaCl下污泥的产甲烷活性.电子传递系统活性(ETS)通过INT〔2-(对碘苯基)-3-(对硝基苯基)-5-苯基氯化四唑〕方法测定〔11〕,具体操作参考文献〔12〕. ...
Enhancing anaerobic digestion and methane production of tetracycline wastewater in EGSB reactor with GAC/NZVI mediator
2
2018
... 以乙酸钠(按COD计,2 000 mg/L)和H2/CO2(140 mL/35 mL)为底物,分别测定污泥的嗜乙酸和嗜氢产甲烷活性.污泥质量与底物COD的比值为1∶1,营养元素组成参考文献〔10〕.用高纯度氮气(99.99%)吹洗血清瓶约1~2 min,立即密封,在35 ℃、150 r/min的摇床中培养,根据甲烷产量随时间变化情况计算比产甲烷活性(SMA).按照上述方法测定0、5、10、15、20 g/L NaCl下污泥的产甲烷活性.电子传递系统活性(ETS)通过INT〔2-(对碘苯基)-3-(对硝基苯基)-5-苯基氯化四唑〕方法测定〔11〕,具体操作参考文献〔12〕. ...
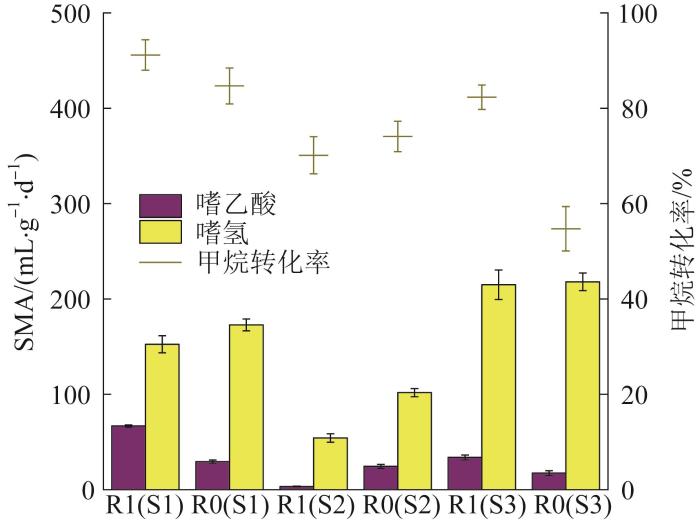
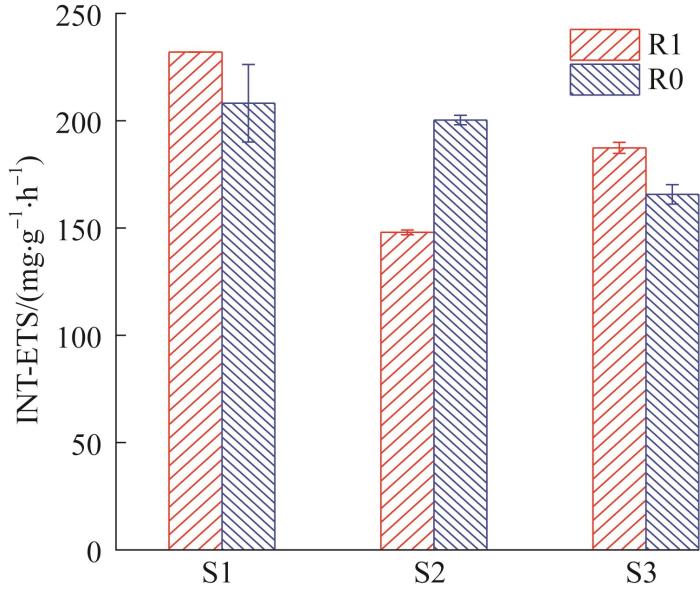
... 污泥的ETS是指污泥微生物呼吸链中的电子传递速率,可间接表征污泥的生物活性.根据Zhaohan ZHANG等〔11〕的研究,INT是评价厌氧微生物系统中ETS活性的较为合适的方法.各阶段R0和R1污泥的INT-ETS活性见图5. ...
Clarifying electron transfer and metagenomic analysis of microbial community in the methane production process with the addition of ferroferric oxide
1
2018
... 以乙酸钠(按COD计,2 000 mg/L)和H2/CO2(140 mL/35 mL)为底物,分别测定污泥的嗜乙酸和嗜氢产甲烷活性.污泥质量与底物COD的比值为1∶1,营养元素组成参考文献〔10〕.用高纯度氮气(99.99%)吹洗血清瓶约1~2 min,立即密封,在35 ℃、150 r/min的摇床中培养,根据甲烷产量随时间变化情况计算比产甲烷活性(SMA).按照上述方法测定0、5、10、15、20 g/L NaCl下污泥的产甲烷活性.电子传递系统活性(ETS)通过INT〔2-(对碘苯基)-3-(对硝基苯基)-5-苯基氯化四唑〕方法测定〔11〕,具体操作参考文献〔12〕. ...
1
2002
... COD、MLSS、MLVSS采用标准方法测定〔13〕.pH采用SG68pH计(METTLER TOLEDO)测定.采用SP-6890气相色谱法(山东瑞宏有限公司)测定沼气中甲烷含量.挥发性脂肪酸(VFAs)采用7890A气相色谱仪(美国 Agilent Technologies)测定. ...
Adsorption and bioadsorption of granular activated carbon (GAC) for dissolved organic carbon (DOC) removal in wastewater
1
2008
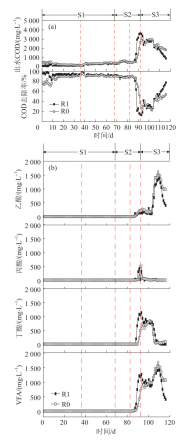
... 启动阶段(S1,第1天至第68天),R1的初始COD去除率超过95%,而R0仅为75%左右,R1的启动时间明显要短于R0〔见图2(a)〕.GAC对部分有机物具有吸附作用,有研究表明GAC可以在90 min内达到吸附饱和〔14〕,而在实际厌氧反应器的环境中,温度和微生物等因素可能会延缓GAC的吸附饱和时间;另一方面,GAC的吸附作用会缓解BOW的生物毒性,提高厌氧微生物的适应性,也可能是启动时间缩短的原因.反应器稳定后(第11天—第36天),R1和R0的COD去除率分别为(93.1±1.0)%和(90.5±1.5)%.当进水COD增加到4 000 mg/L后(第37天—第68天),R1和R0的COD去除率均保持在91%左右.由表2可见,第37天—第68天,R1组甲烷转化率要高于R0组的甲烷转化率.经过长时间的启动驯化,反应器内均形成了降解BOW的微生物群落,这也导致R1和R0在COD去除率方面的差异越来越小.在产甲烷性能上R1依然要优于R0,可能GAC的加入改变了微生物群落结构和电子传递机制,从而增强了厌氧消化产甲烷的效果〔15〕. ...
Granular activated carbon stimulated microbial physiological changes for enhanced anaerobic digestion of municipal sewage
1
2020
... 启动阶段(S1,第1天至第68天),R1的初始COD去除率超过95%,而R0仅为75%左右,R1的启动时间明显要短于R0〔见图2(a)〕.GAC对部分有机物具有吸附作用,有研究表明GAC可以在90 min内达到吸附饱和〔14〕,而在实际厌氧反应器的环境中,温度和微生物等因素可能会延缓GAC的吸附饱和时间;另一方面,GAC的吸附作用会缓解BOW的生物毒性,提高厌氧微生物的适应性,也可能是启动时间缩短的原因.反应器稳定后(第11天—第36天),R1和R0的COD去除率分别为(93.1±1.0)%和(90.5±1.5)%.当进水COD增加到4 000 mg/L后(第37天—第68天),R1和R0的COD去除率均保持在91%左右.由表2可见,第37天—第68天,R1组甲烷转化率要高于R0组的甲烷转化率.经过长时间的启动驯化,反应器内均形成了降解BOW的微生物群落,这也导致R1和R0在COD去除率方面的差异越来越小.在产甲烷性能上R1依然要优于R0,可能GAC的加入改变了微生物群落结构和电子传递机制,从而增强了厌氧消化产甲烷的效果〔15〕. ...
Biofilm formation and granule properties in anaerobic digestion at high salinity
1
2017
... 在恢复阶段(S3,第93天至第116天),去除NaCl后R0和R1的处理效能逐渐恢复.如图2(a)所示,恢复24 d后,R1的COD去除率达到77.2%,而R0仅为53.8%.其间R1和R0的平均甲烷转化率分别为(82.3±2.6)%、(54.7±4.6)%.从图2(b)可以看出,丁酸的降解开始恢复,乙酸开始积累.由于产甲烷菌比产酸菌更加敏感和脆弱〔16〕,产甲烷阶段的恢复滞后,但R1的恢复过程明显快于R0.恢复24 d后,R1中的乙酸质量浓度降至485.95 mg/L,R0为983.10 mg/L,R1的VFA降解速率更快,这与B.CHOWDHURY等〔17〕的研究报道一致. ...
Enhanced biomethane recovery from fat,oil,and grease through co-digestion with food waste and addition of conductive materials
1
2019
... 在恢复阶段(S3,第93天至第116天),去除NaCl后R0和R1的处理效能逐渐恢复.如图2(a)所示,恢复24 d后,R1的COD去除率达到77.2%,而R0仅为53.8%.其间R1和R0的平均甲烷转化率分别为(82.3±2.6)%、(54.7±4.6)%.从图2(b)可以看出,丁酸的降解开始恢复,乙酸开始积累.由于产甲烷菌比产酸菌更加敏感和脆弱〔16〕,产甲烷阶段的恢复滞后,但R1的恢复过程明显快于R0.恢复24 d后,R1中的乙酸质量浓度降至485.95 mg/L,R0为983.10 mg/L,R1的VFA降解速率更快,这与B.CHOWDHURY等〔17〕的研究报道一致. ...
Enhanced anaerobic phenol degradation by conductive materials via EPS and microbial community alteration
1
2018
... 由图5可见,在S1和S3阶段,与R0相比,R1的INT-ETS活性分别提高16.4%、13.1%.Wangwang YAN等〔18〕在处理含酚废水时添加碳纳米管,使污泥的INT-ETS活性提高了10倍,并将这种变化归因于互营菌的富集.因此,本研究中INT-ETS增加很可能是由于GAC对相关互营微生物的富集作用.在S2阶段,R1的INT-ETS活性为(137.6±18.0) mg/(g·h),受高盐度影响明显下降,而R0组为(200.3±2.2) mg/(g·h),变化不大.可见R1的INT-ETS活性更易受盐度影响.结合2.2中盐度对产甲烷活性的影响,认为可能原因为:(1)添加GAC后,微生物之间存在新的电子传递途径(如DIET,而不是传统的IET),高盐环境对其影响更大;(2)在R1中起关键作用的相关微生物因耐盐性差而受到抑制,导致厌氧系统受到显著影响. ...
Enhancing syntrophic associations among Clostridium butyricum,Syntrophomonas and two types of methanogen by zero valent iron in an anaerobic assay with a high organic loading
2
2018
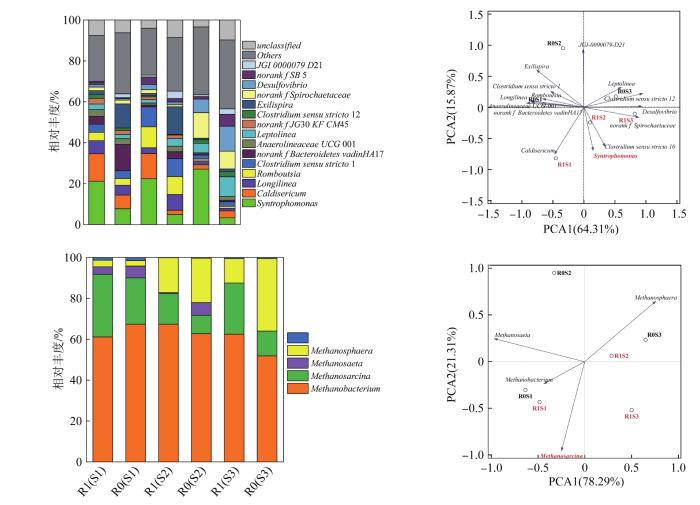
... 反应器的宏观性能主要取决于微观的微生物群落结构,因此研究不同环境(驯化、盐冲击和酸化)中微生物的群落,对阐述GAC-BAR工艺处理效能的提升至关重要.微生物的群落分析如图6所示.由图6(a)可见,R1中各阶段Syntrophomonas的相对丰度(21.2%、22.4%和27.1%)较R0的分别提高了174%、354%、716%,说明Syntrophomonas在R1中明显富集.而Syntrophomonas是一种互营丁酸氧化菌〔19〕,不仅可通过β-氧化途径参与丁酸的降解,还与产甲烷菌相互作用,提高厌氧消化性能.有研究在补充导电材料的复杂环境中检测到Syntrophomonas〔19-20〕.Huijuan LI等〔20〕在水稻土中添加磁铁矿,Syntrophomonadaceae和Methanosarcinaceae的丰度显著增加,促进了丁酸向甲烷的代谢作用.Wei ZHANG等〔21〕在以长链脂肪酸为主的菜籽油中添加GAC,富集了Syntrophomonas和Methanosarcina,显著提高了甲烷产量,缓解了酸化现象.与Syntrophomonas互营共生的古菌为嗜氢产甲烷菌Methanobacterium和嗜乙酸产甲烷菌Methanosarcina.图6(b)中,Methanobacterium是绝对的优势古菌,这与BOW厌氧产甲烷以嗜氢产甲烷途径为主一致.而另一种优势菌Methanosarcina的变化更值得关注.在盐度冲击(S2)下,Methanosarcina(30.7%、15.0%和25.0%)有被Methanosphaera(3.3%、17.0%和11.9%)取代的趋势.但盐度去除后(S3)Methanosarcina的相对丰度得到恢复.且在整个实验过程中,R1的相对丰度比R0增加8.0%~12.8%.因此,GAC的加入很有可能促进了Syntrophomonas与Methanosarcina的互营共生.这从优势菌的分布图〔图6(c)、(d)〕可以清楚地看到,各阶段中R1最核心的菌属是Syntrophomonas和Methanosarcina. ...
... 〔19-20〕.Huijuan LI等〔20〕在水稻土中添加磁铁矿,Syntrophomonadaceae和Methanosarcinaceae的丰度显著增加,促进了丁酸向甲烷的代谢作用.Wei ZHANG等〔21〕在以长链脂肪酸为主的菜籽油中添加GAC,富集了Syntrophomonas和Methanosarcina,显著提高了甲烷产量,缓解了酸化现象.与Syntrophomonas互营共生的古菌为嗜氢产甲烷菌Methanobacterium和嗜乙酸产甲烷菌Methanosarcina.图6(b)中,Methanobacterium是绝对的优势古菌,这与BOW厌氧产甲烷以嗜氢产甲烷途径为主一致.而另一种优势菌Methanosarcina的变化更值得关注.在盐度冲击(S2)下,Methanosarcina(30.7%、15.0%和25.0%)有被Methanosphaera(3.3%、17.0%和11.9%)取代的趋势.但盐度去除后(S3)Methanosarcina的相对丰度得到恢复.且在整个实验过程中,R1的相对丰度比R0增加8.0%~12.8%.因此,GAC的加入很有可能促进了Syntrophomonas与Methanosarcina的互营共生.这从优势菌的分布图〔图6(c)、(d)〕可以清楚地看到,各阶段中R1最核心的菌属是Syntrophomonas和Methanosarcina. ...
Direct inter species electron transfer accelerates syntrophic oxidation of butyrate in paddy soil enrichments
2
2015
... 反应器的宏观性能主要取决于微观的微生物群落结构,因此研究不同环境(驯化、盐冲击和酸化)中微生物的群落,对阐述GAC-BAR工艺处理效能的提升至关重要.微生物的群落分析如图6所示.由图6(a)可见,R1中各阶段Syntrophomonas的相对丰度(21.2%、22.4%和27.1%)较R0的分别提高了174%、354%、716%,说明Syntrophomonas在R1中明显富集.而Syntrophomonas是一种互营丁酸氧化菌〔19〕,不仅可通过β-氧化途径参与丁酸的降解,还与产甲烷菌相互作用,提高厌氧消化性能.有研究在补充导电材料的复杂环境中检测到Syntrophomonas〔19-20〕.Huijuan LI等〔20〕在水稻土中添加磁铁矿,Syntrophomonadaceae和Methanosarcinaceae的丰度显著增加,促进了丁酸向甲烷的代谢作用.Wei ZHANG等〔21〕在以长链脂肪酸为主的菜籽油中添加GAC,富集了Syntrophomonas和Methanosarcina,显著提高了甲烷产量,缓解了酸化现象.与Syntrophomonas互营共生的古菌为嗜氢产甲烷菌Methanobacterium和嗜乙酸产甲烷菌Methanosarcina.图6(b)中,Methanobacterium是绝对的优势古菌,这与BOW厌氧产甲烷以嗜氢产甲烷途径为主一致.而另一种优势菌Methanosarcina的变化更值得关注.在盐度冲击(S2)下,Methanosarcina(30.7%、15.0%和25.0%)有被Methanosphaera(3.3%、17.0%和11.9%)取代的趋势.但盐度去除后(S3)Methanosarcina的相对丰度得到恢复.且在整个实验过程中,R1的相对丰度比R0增加8.0%~12.8%.因此,GAC的加入很有可能促进了Syntrophomonas与Methanosarcina的互营共生.这从优势菌的分布图〔图6(c)、(d)〕可以清楚地看到,各阶段中R1最核心的菌属是Syntrophomonas和Methanosarcina. ...
... 〔20〕在水稻土中添加磁铁矿,Syntrophomonadaceae和Methanosarcinaceae的丰度显著增加,促进了丁酸向甲烷的代谢作用.Wei ZHANG等〔21〕在以长链脂肪酸为主的菜籽油中添加GAC,富集了Syntrophomonas和Methanosarcina,显著提高了甲烷产量,缓解了酸化现象.与Syntrophomonas互营共生的古菌为嗜氢产甲烷菌Methanobacterium和嗜乙酸产甲烷菌Methanosarcina.图6(b)中,Methanobacterium是绝对的优势古菌,这与BOW厌氧产甲烷以嗜氢产甲烷途径为主一致.而另一种优势菌Methanosarcina的变化更值得关注.在盐度冲击(S2)下,Methanosarcina(30.7%、15.0%和25.0%)有被Methanosphaera(3.3%、17.0%和11.9%)取代的趋势.但盐度去除后(S3)Methanosarcina的相对丰度得到恢复.且在整个实验过程中,R1的相对丰度比R0增加8.0%~12.8%.因此,GAC的加入很有可能促进了Syntrophomonas与Methanosarcina的互营共生.这从优势菌的分布图〔图6(c)、(d)〕可以清楚地看到,各阶段中R1最核心的菌属是Syntrophomonas和Methanosarcina. ...
Free acetic acid as the key factor for the inhibition of hydrogenotrophic methanogenesis in mesophilic mixed culture fermentation
1
2018
... 反应器的宏观性能主要取决于微观的微生物群落结构,因此研究不同环境(驯化、盐冲击和酸化)中微生物的群落,对阐述GAC-BAR工艺处理效能的提升至关重要.微生物的群落分析如图6所示.由图6(a)可见,R1中各阶段Syntrophomonas的相对丰度(21.2%、22.4%和27.1%)较R0的分别提高了174%、354%、716%,说明Syntrophomonas在R1中明显富集.而Syntrophomonas是一种互营丁酸氧化菌〔19〕,不仅可通过β-氧化途径参与丁酸的降解,还与产甲烷菌相互作用,提高厌氧消化性能.有研究在补充导电材料的复杂环境中检测到Syntrophomonas〔19-20〕.Huijuan LI等〔20〕在水稻土中添加磁铁矿,Syntrophomonadaceae和Methanosarcinaceae的丰度显著增加,促进了丁酸向甲烷的代谢作用.Wei ZHANG等〔21〕在以长链脂肪酸为主的菜籽油中添加GAC,富集了Syntrophomonas和Methanosarcina,显著提高了甲烷产量,缓解了酸化现象.与Syntrophomonas互营共生的古菌为嗜氢产甲烷菌Methanobacterium和嗜乙酸产甲烷菌Methanosarcina.图6(b)中,Methanobacterium是绝对的优势古菌,这与BOW厌氧产甲烷以嗜氢产甲烷途径为主一致.而另一种优势菌Methanosarcina的变化更值得关注.在盐度冲击(S2)下,Methanosarcina(30.7%、15.0%和25.0%)有被Methanosphaera(3.3%、17.0%和11.9%)取代的趋势.但盐度去除后(S3)Methanosarcina的相对丰度得到恢复.且在整个实验过程中,R1的相对丰度比R0增加8.0%~12.8%.因此,GAC的加入很有可能促进了Syntrophomonas与Methanosarcina的互营共生.这从优势菌的分布图〔图6(c)、(d)〕可以清楚地看到,各阶段中R1最核心的菌属是Syntrophomonas和Methanosarcina. ...
Changes in composition and microbial communities in excess sludge after heat-alkaline treatment and acclimation
1
2010
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Response of fermenting bacterial and methanogenic archaeal communities in paddy soil to progressing rice straw degradation
1
2018
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Evidence of syntrophic acetate oxidation by spirochaetes during anaerobic methane production
1
2015
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Long-term performance and metagenomic analysis of full-scale anaerobic granular sludge bioreactors for low aerobically-biodegradable synthetic fiber manufacturing wastewater treatment
1
2020
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Granular activated carbon alleviates the combined stress of ammonia and adverse temperature conditions during dry anaerobic digestion of swine manure
1
2021
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Digested sludge-degrading and hydrogen-producing bacterial floras and their potential for biohydrogen production
1
2017
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Functional profiling of bacterial communities in Lake Tuz using 16S rRNA gene sequences
1
2021
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Using DNA-based stable isotope probing to reveal novel propionate-and acetate-oxidizing bacteria in propionate-fed mesophilic anaerobic chemostats
1
2019
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Unveiling the mechanisms of medium-chain fatty acid production from waste activated sludge alkaline fermentation liquor through physiological,thermodynamic and metagenomic investigations
0
2020
New insights into the effect of ethanol and volatile fatty acids proportions on methanogenic activities and pathways
1
2021
... 对于R0,由于受盐度和酸积累的影响,其微生物群落从启动阶段开始经历了相似的生态演替.在S1阶段,R0的核心细菌属(相对丰度>8%)为norank f Bacteroidetes vadinHA17(12.9%)和Exilispira(11.8%).有报道称,norank f Bacteroidetes vadinHA17将葡萄糖降解为乙酸、丙酸和H2/CO2〔22〕,与氢营养型产甲烷菌密切相关〔23〕.Exilispira与氢营养产甲烷菌一起参与厌氧反应器中互营乙酸的氧化〔24-25〕.在S2阶段,R0核心菌属分别为Exilispira(13.6%)、Romboutsia(8.8%)和Clostridium sensu stricto 1(8.8%).Clostridium sensu stricto 1是一种常见的产氢细菌属,对不利环境有较强的耐受性〔26〕.Romboutsia是一种与水解和酸化有关的细菌属,尤其与乙酸生成和嗜氢产甲烷有关〔27〕.Romboutsia和Clostridium Sensu Stricto 1相对丰度的增加可能与耐盐性有关.H. A. OYEWISI等〔28〕利用16S rRNA基因序列分析高盐湖细菌群落功能时发现,Romboutsia和Clostridium Sensu Stricto 1是最具代表性的细菌属.在S3阶段R0的核心细菌属分别为Desulfovibrio(12.2%)、Leptolinea(9.6%)和norank f Spirochaetaceae(8.7%).这些优势菌通常在发酵液中富集并降解丙酸〔29-31〕,在酸化环境中相对丰度明显增加.可以看出,R0的核心细菌都是常见的水解酸化细菌,这些优势细菌的具体功能与嗜氢产甲烷菌密切相关〔如图6(b)所示〕.专性氢营养型甲烷菌Methanobacterium占绝对优势(65.0%、61.4%和51.8%).古菌演替的主要变化菌属是Methanosphaera和Methanosarcina.Methanosphaera(2.7%、21.7%和35.4%)逐渐取代Methanosarcina(22.7%、9.0%和12.2%)的优势地位.这说明在R0中难以确定产酸菌和产甲烷古菌的共生关系. ...
Anaerobic biological treatment of industrial saline wastewater:Fixed bed reactor performance and analysis of the microbial community structure and abundance
1
2020
... 由表3可以看出,R1的细菌多样性(Shannon 3.60、3.78和3.68)要低于R0(3.84、4.32和4.02),丰富度(Chao 1)则相反,R1的细菌丰富度高于R0(445.11、593.25和449.34).说明添加GAC促进了特定功能微生物的富集,并且降低了微生物多样性.Syntrophomonas与Methanosarcina的富集可能会导致环境抵抗性能下降:在高盐环境下(S2),Syntrophomonas仍为优势菌,说明Syntrophomonas具有较强的耐盐性(Syntrophomonas是Firmicutes的一员,在之前的研究中,Firmicutes是高盐环境中的优势菌门〔32〕).Methanosarcina相对丰度下降表明其耐盐性较差(在驯化高钠食物垃圾处理过程中,Methanosaricina几乎被嗜氢产甲烷菌Methanobacterium和Methanocpusculum取代〔33〕).因此,Methanosarcina的不耐盐性可能是R1受到盐冲击后处理效能下降的主要原因. ...
The evolution of microbial community during acclimation for high sodium food waste anaerobic digestion
1
2020
... 由表3可以看出,R1的细菌多样性(Shannon 3.60、3.78和3.68)要低于R0(3.84、4.32和4.02),丰富度(Chao 1)则相反,R1的细菌丰富度高于R0(445.11、593.25和449.34).说明添加GAC促进了特定功能微生物的富集,并且降低了微生物多样性.Syntrophomonas与Methanosarcina的富集可能会导致环境抵抗性能下降:在高盐环境下(S2),Syntrophomonas仍为优势菌,说明Syntrophomonas具有较强的耐盐性(Syntrophomonas是Firmicutes的一员,在之前的研究中,Firmicutes是高盐环境中的优势菌门〔32〕).Methanosarcina相对丰度下降表明其耐盐性较差(在驯化高钠食物垃圾处理过程中,Methanosaricina几乎被嗜氢产甲烷菌Methanobacterium和Methanocpusculum取代〔33〕).因此,Methanosarcina的不耐盐性可能是R1受到盐冲击后处理效能下降的主要原因. ...





 津公网安备 12010602120337号
津公网安备 12010602120337号{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}