


随着我国工业的不断发展,工业废水的处理需求日益增加。目前工业废水的处理方法主要有生物法、化学法和物理法3种,其中絮凝法因简单、方便、高效的特点广受青睐。絮凝剂主要分为以下3种:(1)无机絮凝剂,如明矾、聚氯化铝;(2)合成有机絮凝剂,如聚丙烯酰胺衍生物、聚乙烯亚胺;(3)天然或自然产生的高分子絮凝剂,如生物絮凝剂、微生物絮凝剂。无机絮凝剂和合成有机絮凝剂絮凝性能良好稳定、成本低廉,是目前工业废水处理中最常用的絮凝剂。但是这2种絮凝剂的使用也存在一些问题,如无机絮凝剂的大量投加可能会造成活性污泥中金属盐(Fe、Al)的积累,增加后续污泥处理的难度〔1〕。同时无机絮凝剂对废水pH要求较为苛刻,限制了其广泛使用〔2〕。合成有机絮凝剂如聚丙烯酰胺,虽然本身无毒,但在特殊条件下会发生缓慢降解,降解后产生的丙烯酰胺单体毒性很大,对水环境乃至人体健康存在危害〔3〕。微生物絮凝剂一般是指由微生物本身或者从微生物中提取的多糖、蛋白质、脂质、核酸等物质通过纯化加工得到的一类生物制剂〔4〕,其具有处理多种工业废水的潜力,如重金属废水、染料废水、食品加工废水、石化废水等〔5-8〕。微生物絮凝剂同无机絮凝剂及合成有机絮凝剂相比,具有适应性强、可生物降解、无二次污染的优点,具有良好的应用前景。
笔者主要对微生物絮凝剂的研究动态及其在工业水处理中的应用探索进行总结,并对微生物絮凝剂的发展趋势进行预测,以期为确定微生物絮凝剂后续研究方向提供一定参考。
1 微生物絮凝剂的制备与表征
1.1 来源与制备
与传统絮凝方法相比,微生物絮凝剂安全性更高,来源更加广泛。微生物絮凝剂的来源见表1。
表1 微生物絮凝剂的来源
Table 1
| 菌种来源 | 菌种 | 絮凝剂活性组分 | 作用对象 | 参考文献 |
|---|---|---|---|---|
| 细菌 | Brevundimonas,Moraxclla,Psudomonus | 胞外多糖、脂多糖 | 浊度、悬浮物 | 〔9〕 |
| Aliiglaciecola lipolytica | 蛋白质、腐殖酸 | 偶氮染料 | 〔10〕 | |
| Citrobacter W4 | 蛋白质 | 微藻 | 〔11〕 | |
| Bacillus sp. | 多糖 | 重金属离子 | 〔12〕 | |
| Klebsiella variicola B16 | 多糖、蛋白质 | 浊度、悬浮物 | 〔13〕 | |
| Klebsiella pneumoniae strain NJ7 | 多糖、蛋白质 | COD、浊度 | 〔14〕 | |
| Bacillus cereus strain MSI021 | 多糖 | 重金属 | 〔15〕 | |
| Bacillus megaterium strain PL8 | 多糖、氨基酸 | 色度、Pb2+ | 〔16〕 | |
| Raoultella ornithinolytica 160-1 | 多糖、蛋白质 | COD、浊度、色度 | 〔17〕 | |
| Diaphorobacter nitroreducens R9 | 多糖、蛋白质 | COD、浊度、木质素 | 〔18〕 | |
| 真菌 | Phanerochaete chrysosporium BKMF-1767 | 多糖、蛋白质、类氨基糖化合物 | COD | 〔19〕 |
| Aspergillus oryzae(No. CGMCC 3.16041) | 蛋白质 | 浊度 | 〔20〕 | |
| 复合菌种 | Aspergillus fumigatus,Synechocystis sp. PCC6803 | 胞外多糖、胞外蛋白 | Cd、微藻 | 〔21〕 |
| Enterococcus faecalis(MG996001),Proteus mirabilis(MG996004),Lysini bacillus sp.(MG996000) | 多糖 | 重金属离子、蚊虫幼虫 | 〔22〕 | |
| 放线菌 | Streptomyces sp. | 多糖 | 微藻 | 〔23〕 |
| Nocardiopsis dassonvillei(MH900216) | 多糖、脂质 | 浊度 | 〔24〕 | |
| Nocardiopsis sp. GRG 3(KT235642) | 多糖、蛋白质 | 重金属离子 | 〔25〕 |
由表1可看出,微生物絮凝剂主要包括细菌絮凝剂、真菌絮凝剂、复合菌种絮凝剂、微藻絮凝剂等,与微生物的代谢活动密切相关。微生物絮凝剂的主要成分为多糖、蛋白质和脂质等物质,可用于絮凝沉淀工业污水中的重金属离子、悬浮物(SS)、COD等污染物质。
微生物絮凝剂的菌种来源一般为污染场地。Xiang XIA等〔13〕从土壤和活性污泥样品中成功分 离并鉴定了生产絮凝剂MBF-B16的克雷伯氏菌B16,P. SIVASANKAR等〔23〕从河口红树林沉积物中成功分离并鉴定了可用于微藻回收絮凝剂生产的放线菌Streptomyces sp.,均体现了从污染场地分离目标菌种的高效性。基因工程技术也被用于赋予或加强特定菌种的絮凝能力。S. MALIK等〔26〕阐述了将基因修饰应用于微藻絮凝剂生产和通过微藻细胞絮凝来实现高效捕获的可能性。E. DÍAZ-SANTOS等〔27〕通过研究发现,与非转化体相比,贝酵母紫外变种(Saccharomyces bayanus var. uvarum)对淡水微藻莱茵衣藻的絮凝效率提高了2~3倍,证实了以基因工程菌作为絮凝剂生产菌种来源的巨大潜力。
微生物絮凝剂的制作过程一般包括筛选、分离、培养、提取、纯化。从污染场地获得待筛选菌株后,需将菌株接种于特定的培养基中进行筛选。Jingqiu HUA等〔12〕在获得Bacillus sp.菌株后,将其置于M9培养基中培养,待菌株培养至预设OD值后,再通过离心和超声处理的方法将絮凝物质与菌株分离,最终通过离心、乙醇萃取以及透析和冷冻干燥收集纯化后的生物絮凝剂。
1.2 表征
微生物絮凝剂的研究通常是按照由表及里的逻辑开展的。以此为基础,微生物絮凝剂的表征方法可分为2类,一类用于絮凝剂表层结构观测及絮凝能力初步评估,如扫描电子显微镜(SEM)、场发射扫描电子显微镜(FE-SEM)、环境扫描电子显微镜(ESEM)等;另一类用于深度研究絮凝剂的稳定性及其组成、结构和絮凝能力,如热重分析(TG)、示差扫描量热法(DSC)、Zeta电位、傅里叶变换红外(FT-IR)光谱、气相色谱-质谱联用(GC-MS)、能量色散X射线光谱(EDX)、X射线光电子能谱(XPS)、三维荧光光谱(3D-EEM)等。常见表征方法及其作用见图1。
图1
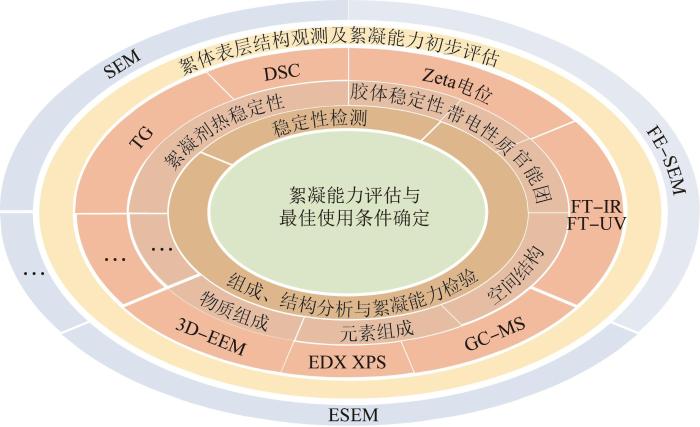
通过SEM及相关技术获得的显微图像是研究絮凝剂表层结构及评估其絮凝能力的重要工具。Junjun WANG等〔21〕采用SEM观测了真菌絮凝捕获微藻的过程,结果显示真菌菌丝颗粒捕获、固定了微藻,并与微藻发生了交联,成功证实了微生物絮凝剂回收微藻的可能性。Yong NIE等〔20〕使用FE-SEM对真菌-高岭土颗粒的表面形态进行了表征,真菌颗粒表面具有多孔褶皱,可作为絮凝剂有效聚集高岭土颗粒,达到降低饮用水中浊度的目的。杨朝晖等〔28〕采用ESEM技术探究了十二烷基苯磺酸钠(SDBS)与Ca2+的作用过程及其结合微生物絮凝剂GA1去除废水中罗丹明B的原理,ESEM图谱显示,随着SDBS投入量的增加,絮体结构愈发紧密且表面愈发粗糙,为絮凝颗粒不断复溶致使絮体更加密实提供了有力证据。
除了对絮凝剂表观结构进行观测外,为进一步研究其作用机制并发掘更大应用潜力,还需对絮凝剂结构、组成和性能(絮凝能力和稳定性)进行更深入的表征。
对絮凝剂稳定性的表征方法有2类,即对其热稳定性进行表征的DSC、TG等技术和对其絮体稳定性进行表征的Zeta电位等。母哲轩等〔29〕筛选出一种可高效产生絮凝剂的巨大芽孢杆菌菌株TF10并采用TG法对其热稳定性进行了表征,结果显示,絮凝剂从210 ℃开始失重;在350 ℃时,失重率达到12%;在700 ℃时,质量仅存1.2%。由此可见,在一般温度条件下,该絮凝剂具有良好的热稳定性。D. K. KARTHIGA等〔30〕用DSC测定了芽孢杆菌生产的絮凝剂的耐热性能,最终发现3种絮凝剂可耐受100~150 ℃的高温。吴鹏等〔31〕通过分析Zeta电位的变化发现,同时投加Fe和微生物絮凝剂时废水的Zeta电位绝对值最小,废水中的胶体更易于脱稳,进而促进沉降。
按照不同层次,对絮凝剂的组成、结构和性能的表征可分为物质组成、元素组成、官能团组成和带电性质等〔32〕。R. T. V. VIMALA等〔22〕用TG法测定了生物絮凝剂的热解产物,结果显示,多糖可能是生物絮凝剂的成分之一,其官能团可作为悬浮物的吸附点,从而提高絮凝效率。Yunxiao WANG等〔10〕采用3D-EEM光谱对胞外聚合物(EPS)样本进行了检测,在不同EPS的光谱中均识别出3个峰,分别为色氨酸、芳香族氨基酸以及腐殖酸。G. RAJIVGANDHI等〔25〕采用XPS分析了生物絮凝剂的原子和元素成分,结果显示,元素C、Na、O、N的摩尔分数分别为39.16%、41.22%、10.48%、1.97%。采用FT-IR等技术可对絮凝剂的官能团进行表征。穆军等〔33〕对细菌Halomonas sp. GHF11产生的絮凝剂进行了FT-IR检测,通过光谱图推测出其中存在羰基、羟基、羧基等基团。Zeta电位可对絮凝剂的带电性质与絮凝机理进行判断。Weihua LI等〔9〕分别测量了北京地区地表水菌群的Zeta电位和电导率,观测到EPS中阴离子含量显著下降,证明了EPS的絮凝机制并非电荷中和,而是架桥和吸附作用。杨志超等〔34〕在研究多糖型微生物絮凝剂对闪锌矿与方解石的絮凝性能过程中进行了Zeta电位分析,絮凝剂在污水中呈负电性,但其对方解石的絮凝作用并非经静电作用实现,而是以化学键合作用为主导。
2 微生物絮凝剂的絮凝机制
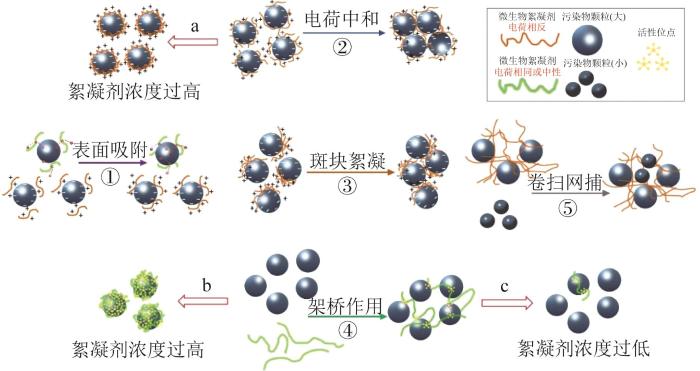
目前公认的微生物絮凝剂的絮凝机制包括表面吸附、电荷中和、斑块絮凝、架桥作用、卷扫网捕以及其他作用等。在微生物絮凝中起主要作用的机制是电荷中和和架桥作用,这可能与微生物絮凝剂存在众多活性基团(羧基、羟基、氨基、亚甲基等)有很大关系。然而,实际过程中的絮凝往往是多种絮凝机制共同作用的结果。因为实际废水的复杂性和特异性,絮凝剂的自身性质(分子质量、结构、电荷密度、用量)和污染物性质(电性、pH、温度、离子强度)都将对絮凝机制产生较大影响。
微生物絮凝剂的主要絮凝机制见图2。
图2
2.1 表面吸附作用
在絮凝剂刚进入溶液时,絮凝剂会在污染物颗粒的表面活性位点发生吸附,这是絮凝得以发生的必要条件。常见的表面吸附包括静电作用和氢键2种(图2紫色箭头①)。与污染物颗粒带相反电荷的絮凝剂在静电引力下能吸附在颗粒表面,有研究发现,带正电的多糖改性絮凝剂对带负电的高岭土颗粒的吸附作用促进了絮凝剂对金属离子的絮凝〔35〕。絮凝剂通常可以通过改性获得更长的分子链和更高的分子质量来提升其表面吸附效果,对絮凝剂进行化学改性以增加其表面的氨基、羟基等官能团数量是优化絮凝剂的一种常用方法。以氢键为作用力的表面吸附通常靠氢原子和电负性原子之间的吸引作用实现,絮凝剂上可能存在的酰胺基、醚氧基等容易与悬浮颗粒上的氢原子形成氢键,使得絮体增大〔36〕。表面吸附过程对胶体稳定性有很大影响,当吸附量较大时,空间位阻稳定现象开始发挥作用〔37〕;当只有部分表面被覆盖时,颗粒可以变成不稳定的吸附聚合物,导致絮凝。
2.2 电荷中和作用
在表面吸附进行一段时间后,对于表面电荷分布均匀的污染物颗粒,电荷中和起主要作用(图2蓝色箭头②)。该机制主要基于扩散双电层模型和DLVO理论,废水中大多数分散的污染物颗粒带同种电荷,颗粒间的静电斥力保证了系统长期稳定。然而,向废水中加入带相反电荷的微生物絮凝剂时,污染物颗粒的双电层被压缩,Zeta电位降低,颗粒间斥力减小,颗粒碰撞时很容易发生絮凝。基于该机制,中和作用一般可通过Zeta电位的变化判断,如果系统的最佳絮凝点处的Zeta电位接近于零,则絮凝主要由电荷中和作用引起〔38〕。但需要注意的是,絮凝剂并非越多越好,在最佳絮凝点往后,随絮凝剂用量进一步增加,污染物颗粒的Zeta电位会逐渐偏离零,实现电荷反转,污染物颗粒出现“再稳定”的状态(图2红色箭头a)。因此,在以电荷中和作用为主的絮凝过程中,通常需要结合Zeta电位的变化曲线确定絮凝剂的最佳投加量。
2.3 斑块絮凝作用
2.4 架桥作用
电荷中和和斑块絮凝作用主要针对絮凝剂和污染物颗粒电性相反的情况。而对于中性或与污染物带同种电荷的絮凝剂,除了向体系中投加金属阳离子加强絮凝剂的吸附和中和作用以外〔40〕,最主要的作用机制是依靠架桥作用实现絮凝。在微生物絮凝剂上的有效附着位点吸附污染物后,链上未被吸附的部分会向外延伸,形同“架桥”,吸附并连接其他更多污染物(图2绿色箭头④)。架桥作用一般依靠范德华力、静电、氢键作用,甚至是多糖分子的自由基与颗粒之间的化学反应实现。通常絮凝剂分子质量越大,其絮凝效果越好,因此很多研究都通过接枝改性来提高絮凝剂的架桥絮凝效果〔41〕。但絮凝剂分子链过长时会产生絮凝剂自身折叠的情况,从而阻碍胶粒的靠近。絮凝剂的用量对架桥作用也很重要,絮凝剂过少,不足以形成架桥作用(图2红色箭头c);絮凝剂过多,颗粒表面被絮凝剂完全“包覆”,导致架桥作用受阻,影响絮凝的发生(图2红色箭头b)。因此有研究对絮凝剂最佳用量进行了分析,V. RUNKANA等〔42〕建立了关于架桥絮凝的数学模型,发现絮凝剂用量约为饱和量的50%时,絮凝效果最好。
2.5 卷扫网捕作用
网捕作用一般发生在中和作用和架桥作用之后,较大的絮凝体逐渐形成,这些大的絮凝体在运动或者沉降时会卷入或者捕集一些较小的污染物颗粒,然后形成更大的絮凝体(图2灰色箭头⑤)。污染物浓度对该过程有较大影响,通常污染物颗粒浓度越高,卷扫网捕作用越强。
2.6 其他作用
由于微生物絮凝剂的链骨架通常含有许多活性官能团,它们能够通过一些高度特异的力与污染物上相应的官能团发生化学反应或结合,如疏水链与疏水污染物之间的疏水相互作用〔43〕、重金属和絮凝剂官能团的配位及螯合作用、芳香环基微生物絮凝剂和芳香族污染物的π-π叠加作用。
3 微生物絮凝剂生产使用的优化
3.1 物化条件
物化条件的优化对于絮凝剂的生产使用至关重要,目前的大部分研究从微生物的培养条件(碳氮源、C/N、温度、pH、培养时间等)和絮凝剂使用条件(絮凝剂用量、阳离子、pH、温度等)对多种物化因子进行了优化,常用的优化分析方法有单因素法、正交试验法、响应曲面法等。
3.1.1 碳氮源
絮凝剂生产中常用的碳源包括葡萄糖、淀粉、蔗糖等,常用的氮源包括尿素、酵母粉、牛肉膏、蛋白胨等。微生物对利用底物存在特异亲和性,不同微生物的最佳碳氮源往往不同,产生的絮凝剂结构也有所差异。Changqing ZHAO等〔44〕通过单因素实验证明20 g/L的葡萄糖是枯草芽孢杆菌CZ1003生产絮凝剂的最佳碳源;而在B. A. RASULOV等〔45〕的实验中,相比葡萄糖,D-甘露糖更适合作为生产根瘤菌SZ4S7S14絮凝剂的碳源。Ou LI等〔46〕发现蛋白胨对埃吉类芽孢杆菌B69生产絮凝剂非常有利,而黄曲霉生产絮凝剂的最佳氮源是酵母膏和尿素〔47〕。某些研究表明,相比单一碳源,复合碳源更有助于提高絮凝剂的产量和质量。V. AJAO等〔48〕以甘油和乙醇作为污水处理厂好氧污泥的碳源,相比单一碳源培养时,絮凝剂的产量和絮凝效率均有所提高。微生物絮凝剂的生产成本历来是广大研究者关注的问题,利用现有的废弃物和废水作为培养基质不仅可以降低生产成本,还能实现废弃物的资源化利用。玉米秸秆、花生壳、米糠等木质纤维素类废弃物是潜在的碳源,研究者们致力于寻找一种既能分解木制纤维素又能产生絮凝剂的双功能菌株。Weijie LIU等〔49〕发现纤维素分解菌L804可以分泌纤维素酶和木聚糖酶,将秸秆等生物质直接转变成生物絮凝剂,该絮凝剂对微藻的絮凝效率达到99.04%。还有研究者发现硝基还原黄杆菌R9可分泌木质素酶和纤维素酶,将苎麻生物质转化为絮凝剂〔18〕。高浓度的有机废水同样是理想的替代碳源,如马铃薯废水、啤酒废水、乳制品废水、畜禽废水、淀粉加工废水等〔14,50-53〕。近年来关于餐厨垃圾、剩余污泥等废弃物用作培养基替代碳源的研究也越来越多〔49,54-55〕。相比碳源的优化,目前关于低成本氮源的研究较少。据研究,以羽毛废弃物为嗜碱琼脂芽孢杆菌C9廉价替代氮源生产的生物絮凝剂可用于处理草浆洗涤废水〔56〕。也有研究者尝试将动物内脏用作培养基的氮源〔57〕,这些研究都在一定程度上解决了絮凝剂的高成本问题。
3.1.2 C/N
C/N对絮凝剂的产量也有重要影响。有研究发现絮凝剂的生产和细胞生长存在底物竞争关系,低C/N有助于提高微生物的生物量,高C/N有助于提高絮凝剂的产量。这可能是在氮源受限时,碳源过剩,细菌可以通过自主调节碳和氮代谢流动的方向,使多余的能量转化为EPS,特别是多糖,并将其作为细菌的碳源和储能物质〔48〕。所以某些情况下,改变C/N可以作为提高絮凝剂产量的一个手段。
3.1.3 温度和pH
培养过程中的温度主要是通过影响相关酶的活性来影响絮凝剂产量,大部分微生物的最佳培养温度在30 ℃左右。絮凝温度同样影响絮凝效果,由于絮凝剂中一般多糖含量高,所以其对温度耐受性较高,如巨型芽孢杆菌生产的絮凝剂在20~80 ℃均能保持90%的絮凝活性〔50〕。但是过高的温度可能会影响生物絮凝剂中蛋白质的结构,造成絮凝剂损伤,降低絮凝效果。同样地,培养基初始pH对絮凝剂产量的影响也不可忽略。pH主要通过影响微生物对营养物质的吸收和细胞酶活性影响絮凝活性。一般最适宜微生物生产絮凝剂的pH为中性或者弱碱性。例如,地衣芽孢杆菌生产的絮凝剂的絮凝活性在生产pH为5~12时始终保持在92%以上,在pH为8时达到最大值97%〔36〕。肺炎克雷伯菌NJ7菌株在pH=10.0时生物絮凝剂的产率有显著提高,且是中性时的2倍〔53〕。絮凝过程中溶液的pH也会影响絮凝效果,很多微生物在较大pH范围内可保持较高的絮凝活性,如蜡样芽孢杆菌在pH 2~10范围内均能保持70%以上的活性〔15〕,这也是微生物絮凝剂可以广泛应用的原因之一。
3.1.4 阳离子
3.2 基因工程
近年来,随着基因组学技术和基因编辑技术的不断发展成熟,利用基因工程的方法优化絮凝剂生产的研究也越来越多。Zhen CHEN等〔59〕通过对地衣芽孢杆菌CGMCC 2876的基因组进行测序,确定了参与细菌EPS合成的eps基因簇。黄单胞杆菌特有絮凝剂黄原胶的合成主要由黄原胶操纵子(包括13个基因)和xanAB基因(前体合成)控制〔60-61〕。琥珀酸是由根瘤菌、农杆菌、假单胞菌等菌株合成的杂多糖,它的生物合成需要19个exo基因和2个exs基因调控〔62〕。基因表达在核苷糖的形成、糖重复单元的组装和聚合以及多糖的转运和出口等过程中发挥了重要作用,因此通过基因工程技术对微生物絮凝剂生产进行调控也成为近年来的热点研究问题。通过基因工程对絮凝剂进行优化的策略主要包括2种,分别是提高微生物絮凝剂的产量和改善微生物絮凝剂的结构。
3.2.1 提高絮凝剂的产量
提高絮凝剂的产量的方法如图3所示,主要有过表达絮凝剂前体合成基因、过表达参与重复单元组装的基因、过表达絮凝剂转运输出的基因以及敲除与絮凝剂合成有竞争关系的基因等方法,最近也有新的研究发现能量或辅因子水平也可能是EPS生物合成的约束。
图3
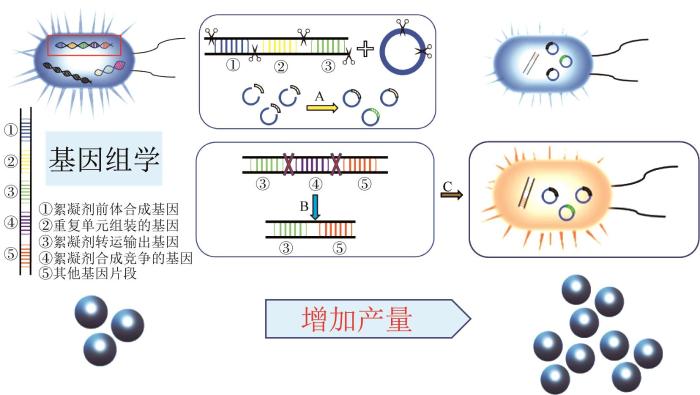
图3
基因工程技术:增加絮凝剂产量
Fig. 3
Genetic engineering technology to increase flocculant production
多糖合成相关基因的过度表达是一种常见方法(图3,A路径,黄色箭头)。Zhen CHEN等〔59〕在地衣芽胞杆菌CGMCC 2876中过表达编码糖基转移酶的epsDEF基因后,重组菌株的絮凝活性比原菌株提高了90%;同样,多糖生物絮凝剂的产量提高了27.8%。D. SENGUPTA等〔63〕过度表达土壤根瘤菌F2中与葡萄糖和半乳糖单体增加有关的exoY基因后,絮凝剂产量提高了14.8%,对六价铬的回收率提高了18%;絮凝剂中葡萄糖和半乳糖的比例增加,而甘露糖和鼠李糖的比例下降。鞘氨醇是一种由鞘氨醇菌属产生的细胞外多糖,Haidong HUANG等〔64〕过度表达了S. sanxanigenens中的pgmG基因(编码磷酸葡萄糖酶和磷酸甘露糖突变酶活性的双功能蛋白),使得鞘氨醇产量增加(17±0.3)%。
3.2.2 改善微生物絮凝剂的结构
随着基因组学和克隆重组技术的发展,通过基因工程对生产絮凝剂特性进行设计的方法将会得到更多关注。
3.3 化学改性
化学改性是指通过化学反应来改变絮凝剂结构的一种优化方法,根据化学反应类型分为接枝共聚反应、醚化反应、胺化反应、酯化反应、酰化反应、氧化反应、交联反应等〔41〕。在众多改性方法中,接枝共聚因操作简单、制备条件温和、产品应用性能好等优势成为化学改性中最常用的方法。
接枝共聚是以絮凝剂为基质,在引发剂作用下,在其分子链上引入具有活性基团的有机聚合物侧链,以达到性能增强的一种技术。引发接枝的方式包括热引发、光引发、辐射引发、微波引发和等离子体引发等〔72〕,常用的接枝单体可分为阴离子型、阳离子型、非离子型3种。在微生物絮凝剂接枝改性中使用较多的阳离子单体主要有(2-甲基丙烯酰氧乙基)三甲基氯化铵(DMC)〔73〕、二甲基二烯丙基氯化铵(DADMAC)〔74〕、2,3-环氧丙基三甲基氯化铵(ETA)〔75〕、甲基丙烯酰胺丙基三甲基氯化铵(MAPTAC)〔76〕、酰氧季铵盐丙烯酰氧乙基三甲基氯化铵(DAC)〔77〕等。阴离子单体主要有丙烯酸(AA)〔78〕,2-丙烯酰胺基-2-甲基丙烷磺酸(AMPS)等〔79〕。非离子单体主要有丙烯酰胺(AM)、苯乙烯、丙烯腈、环氧化物等。近来阴阳离子官能团同时存在的两性接枝单体也引起了研究者们的关注,Lihua LIU等〔80〕采用紫外分光光度法分析了两性螯合聚合物絮凝剂(ACPF),它不仅兼有阴阳离子接枝单体的特性,且适应pH范围广。
接枝共聚发生的部位一般是絮凝剂分子链上的—NH2和—OH,因此拥有丰富—NH2和—OH基团的壳聚糖在絮凝剂改性方面最为常用。Xiaomin TANG等〔79〕以硝酸铈为引发剂,采用接枝共聚法制备了磺化壳聚糖基絮凝剂CS-g-P(AM-AMPS),显著缩短了絮凝所需时间。Yongjun SUN等〔81〕采用紫外光引发接枝聚合法制备了一种新型双功能壳聚糖絮凝剂CMCTS-g-P(AM-CA),该絮凝剂表现出十分优异的絮凝性能。Tao ZENG等〔38〕用微波加热引发接枝反应设计了一种新型葡聚糖基絮凝剂,与传统絮凝剂相比,其架桥和吸附性能增强,絮凝效率得到提高。接枝共聚物的性质主要由其结构特征决定,如接枝比、电荷密度以及连接聚合物链的长度和数量,这些特性受合成条件的影响,特别是总辐照、引发剂剂量和进料单体的量等。因此,对这些条件的考察也是接枝共聚应该关注的问题。
3.4 复合絮凝剂
一些研究发现,将不同种类的絮凝剂进行复合可以显著减少絮凝剂的用量和提升絮凝效果。相比单独使用聚氯化亚铁(PAFC),联合使用微生物絮凝剂(MBF-B16)和PAFC可以降低饮用水中的浊度并去除SS,可节约72%的絮凝剂用量〔13〕。Chaofan ZHANG等〔82〕组合微生物絮凝剂聚γ-谷氨酸和氧化钙来收获普通小球藻,发现该复合絮凝剂主要通过影响微藻细胞的Zeta电位和悬浮液的pH来影响微藻絮凝,其絮凝效率最高达到95%。Z. FERASAT等〔83〕将明矾和酵母细胞壁(YCW)混合使用,可以很大程度缓解加入明矾引起出水pH较低的问题,出水无需再经过pH调节。Jingshen DONG等〔84〕在硅藻土体系中加入壳聚糖,硅藻土的等电点(IEP)发生变化,其对煤浆水的絮凝效率达到84.3%。不同聚合程度的无机絮凝剂也会对复合使用的效果产生影响,Yuanxia LUO等〔85〕研究了不同种类聚合氯化铝(PACl)和浒苔多糖的组合同时去除CuO纳米颗粒、Cu2+和腐殖酸的效果,发现中等聚合度的PAClb对浊度和Cu2+的去除效率最高,高聚合度的PAClc对腐殖酸的去除效果最好,这可能是由污染物的种类不同导致的。还有研究发现,絮凝剂的复合作用可以解决无机絮凝剂过量引起的溶液“再稳定”问题,将AlCl3和生物絮凝剂EPS-160结合使用不仅可以使絮凝效率提高30%,还可使AlCl3的最佳剂量范围变得更广(11~23 mg/L)〔17〕。
3.5 群感效应
4 微生物絮凝剂在工业废水处理中的应用
由于提纯技术要求较高,且提纯后的保存与再溶解均会对絮凝剂的絮凝能力产生较大影响,微生物絮凝剂的生产成本居高不下,尚未实现大规模的工业化生产应用。但微生物絮凝剂在重金属废水、印染废水等工业废水处理中表现出极大的应用潜力,“原位生产,原位使用”的高适应性优势仍吸引着诸多学者对其工业化应用进行探索。
4.1 重金属废水
4.2 印染废水
4.3 其他类型工业废水
微生物絮凝剂在其他类型工业废水处理中也有着广泛的应用前景。Jiawen HE等〔11〕从微藻污水培养系统中分离到柠檬酸杆菌W4,该菌株产生的蛋白质EPS絮凝剂对蛋白核小球藻具有灭活活性,对藻类的回收率达到87.37%,该研究为污水中藻类的去除提供了一种新的技术方法。R. KAUR等〔92〕从堆肥厂渗滤液中分离出一种芽孢杆菌,利用该菌种生产的黏液EPS(S-EPS)和FeSO4混合絮凝处理渗滤液,其对COD、磷、氨氮的去除率分别高达92%、94%、96%。Chunying ZHONG等〔18〕从农业苎麻生物脱胶废水中分离出一种硝基还原黄杆菌R9,该菌可将苎麻生物质转换为絮凝剂MBF-9,在MBF-9用量为831.75 mg/L、pH=7.58以及温度为26.2 ℃的条件下,其对废水中浊度、木质素和COD的最大去除率分别为96.2%、59.2%和79.5%。在微生物絮凝剂广泛应用的过程中,絮凝剂生产菌的菌种来源也有了新的扩充。蝉花作为一种虫生真菌,最初被关注到是因为其独特的药理活性,而Xiao ZOU等〔93〕发现蝉叶板蓝根GZU6722产生的IC-1具有絮凝潜力,对洗煤废水的絮凝效率可达91.84%。
此外,对原有微生物絮凝剂的改进研究也在不断发展。Yizhuo ZHANG等〔94〕以硝酸铈铵作为引发剂,将蜡样芽孢杆菌生产的生物絮凝剂与两性淀粉结合改性,改性絮凝剂的絮凝效率和总氮去除率分别可达98.17%和100.00%,相较于未改性的絮凝剂,絮凝效率和总氮去除率的提升率均超过30%。
表2总结了微生物絮凝剂在工业废水处理中的应用。
表2 微生物絮凝剂在工业废水处理中的应用
Table 2
| 废水来源 | 废水性质 | 絮凝剂来源 | 絮凝剂用量 | 处理效果 | 参考文献 |
|---|---|---|---|---|---|
| 皮革废水 | 高总氮、 高浓度有色物质 | Bacillus cereus | 1 g/L | 絮凝效率98.17%;总氮去除率100.00% | 〔94〕 |
| 堆肥渗滤液 | 高COD、高氮磷、 多种重金属离子 | 从废水污泥中分离出的菌株BS-04 | 0.5 g/L | COD去除率92%;磷去除率94%;氨氮去除率96% | 〔92〕 |
| 印染废水 | 高BOD、高COD、高色度、高浓度重金属离子 | Bacillus subtilis(E1), Exiguobacterium acetylicum(D1), Klebsiella terrigena(R2), Staphylococcus aureus(A22), Pseudomonas pseudoalcaligenes(A17),Pseudomonas plecoglossicida(A14) | 2~10 mg/L | 脱色率97.04% | 〔91〕 |
| 淀粉加工废水 | 高COD、高浊度 | Klebsiella pneumoniae strain NJ7 | 30 mg/L | COD去除率41%;浊度去除率90% | 〔14〕 |
| 秸秆废水 | 高SS | Bacillus agaradhaerens C9 | 6.0 mg/L | SS去除率93.1% | 〔56〕 |
| 制浆废水 | 高COD | Diaphorobacter nitroreducens R9 | 831.57 mg/L | 浊度去除率96.2%;COD去除率79.5%;木质素去除率9.2%;多糖去除率63.1% | 〔18〕 |
| 洗煤废水 | 高浓度SS | Isaria cicadae GZU6722 (IC-1) | 24 mg/L | 最大絮凝效率91.81% | 〔93〕 |
| 高岭土模拟废水 | 高SS | Aspergillus flavus MCB 271 | — | 絮凝活性97%;细菌负荷减少58.73% | 〔95〕 |
| 养猪废水 | 高COD、高TN、高SS | Talaromyces trachyspermus OU5 | 5%(体积分数) | COD去除率52.1%;总氮去除率39.7%; SS去除率75% | 〔96〕 |
5 结语和展望
目前,虽然微生物絮凝剂具有安全无毒、无二次污染、应用范围广等多种优势,但也存在不可忽视的局限性,后续可以从以下几个方面对其进行改善:
(1)目前微生物絮凝剂的使用大部分是“原位生产,原位使用”,产业化难度大。一方面是因为从胶状微生物中提取絮凝剂的难度大、流程复杂,另一方面是因为提取纯化、运输储存过程中絮凝能力的大幅下降影响产品效能。提取絮凝剂常用的有机试剂沉淀法对絮凝剂的活性影响极大,应探索更便宜、温和的提取方法,例如利用溶菌酶对微生物进行酶解,同时结合微波法、超临界萃取法、双水相萃取法等技术实现高效温和提取和提高收获率;在改进用于絮凝剂工业化的机械设备和条件的同时,根据絮凝剂的组成结构优化生产方案,减少不必要的流程;选用廉价培养基(有机废水、废纤维、污泥等),通过基因工程“定制”微生物絮凝剂,合理利用改性、絮凝剂复配等技术降低生产成本,提高絮凝能力。
(2)当前微生物絮凝剂缺乏统一的评价体系和标准,不利于市场管理。因此应尽快建立对其质量和安全性进行评价的体系,逐步制定微生物絮凝剂国家产品标准。
(3)微生物絮凝剂与新兴技术的联合使用也是不容忽视的一点,合成生物学有关技术的发展对推动絮凝剂的开发具有重要意义。通过基因编辑、通路改造、代谢通量优化、酶工程、调控电路重连和宿主修饰等方式对基因模块进行重新编程,无论是对絮凝剂修饰还是生产新型絮凝剂,都是一种可期的方法,这些技术的集成有助于理解复杂的基因调控和代谢网络,实现对基因表达的高效调控。利用人工智能系统快速设计一些絮凝剂中间物合成路线,将其与基因工程技术结合,可实现对絮凝剂生产的准确高效优化。同时,基于算法和现有实验数据建立絮凝剂生产使用的模型,对絮凝剂的使用效果进行预测,可实现絮凝剂生产设计和使用工艺条件的优化,从而极大提高时间效率和降低优化成本。
参考文献
絮凝剂在水处理中的应用与研究进展
[J].
Application and research progress of flocculant in water treatment
[J].
巨大芽孢杆菌多糖型絮凝剂的纯化、结构鉴定和絮凝活性的研究
[D].
Purification,structural characterization and flocculation activity of polysaccharide-based bioflocculants from Bacillus megaterium
[D].
A review of the toxicology of acrylamide
[J].
Extracellular biopolymeric flocculants:Recent trends and biotechnological importance
[J].
絮凝剂处理重金属废水的研究进展
[J].
Research progress in flocculants applied to the treatment of wastewater containing heavy metals
[J].
微生物净化技术在染料废水中的应用分析
[J].
Application analysis of microorganism purification technology in dye waste water
[J].
绿色絮凝剂产生菌的分离及处理啤酒废水研究
[J].
Research on the separation of green flocculant produced strains and the treatment of brewery wastewater
[J].
一株产絮凝剂菌株K8及其絮凝特性研究
[J].
Research on a flocculant-producing strain K8 and its characteristics of flocculation
[J].
Contribution of bacterial extracellular polymeric substances(EPS) in surface water purification
[J].
Treatment of azo dye wastewater by the self-flocculating marine bacterium Aliiglaciecola lipolytica
[J].
A bacterial strain Citrobacter W4 facilitates the bio-flocculation of wastewater cultured microalgae Chlorella pyrenoidosa
[J].
Energy-saving preparation of a bioflocculant under high-salt condition by using strain Bacillus sp. and the interaction mechanism towards heavy metals
[J].
Characterization and coagulation-flocculation performance of a composite flocculant in high-turbidity drinking water treatment
[J].
Recycling of starch processing industrial wastewater as a sole nutrient source for the bioflocculant production
[J].
Revealing the ability of a novel polysaccharide bioflocculant in bioremediation of heavy metals sensed in a Vibrio bioluminescence reporter assay
[J].
Using a novel polysaccharide BM2 produced by Bacillus megaterium strain PL8 as an efficient bioflocculant for wastewater treatment
[J].
Rapid production of a novel Al(Ⅲ) dependent bioflocculant isolated from Raoultella ornithinolytica 160-1 and its application combined with inorganic salts
[J].
Production of a bioflocculant from ramie biodegumming wastewater using a biomass-degrading strain and its application in the treatment of pulping wastewater
[J].
Soluble microbial products from the white-rot fungus Phanerochaete chrysosporium as the bioflocculant for municipal wastewater treatment
[J].
Aspergillus oryzae,a novel eco-friendly fungal bioflocculant for turbid drinking water treatment
[J].
Construction of fungi-microalgae symbiotic system and adsorption study of heavy metal ions
[J].
Characterization of self-assembled bioflocculant from the microbial consortium and its applications
[J].
Characterization of a novel polymeric bioflocculant from marine actinobacterium Streptomyces sp. and its application in recovery of microalgae
[J].
Effective removal of heavy metals in industrial wastewater with novel bioactive catalyst enabling hybrid approach
[J].
Enlightening the characteristics of bioflocculant of endophytic actinomycetes from marine algae and its biosorption of heavy metal removal
[J].
Microalgal flocculation:Global research progress and prospects for algal biorefinery
[J].
Study of bioflocculation induced by Saccharomyces bayanus var. uvarum and flocculating protein factors in microalgae
[J].
基于饱和沉淀Ca(DBS)2及结合MBFGA1絮凝沉降去除罗丹明B
[J].
Removing Rhodamine B based on enhanced precipitation of calcium dodecyl benzene sulfonate combined with microbial flocculation GA1
[J].
微生物絮凝剂絮凝条件的响应曲面法优化
[J].
Optimization of flocculating conditions of microbial flocculant using response surface methodology
[J].
Isolation and characterization of toxic metal removing bacterial bioflocculants
[J].
微生物絮凝剂提取方法的优化及其对废水中木质纤维类污染物的絮凝机理
[J].
Optimization of the extraction method of the bioflocculant and its application in lignocellulosic waste-containing water
[J].
Novel myco-coagulant produced by Lentinus squarrosulus for removal of water turbidity:Fungal identification and flocculant characterization
[J].
一种从菲律宾蛤仔黏附细菌Halomonas sp
GHF11中提取的多糖絮凝剂的表征其在脱色中的应用[J].
Characterization of a novel polysaccharide flocculant extracted from Manila adhesive bacterium Halomonas sp. GHF11 and its application in decolorization
[J].
多糖微生物絮凝剂对方解石与闪锌矿的絮凝作用及机理
[J].
Role and mechanism of polysaccharide-based microbial flocculant in flocculation calcite and sphalerite
[J].
Extracellular polymeric substances of bacteria and their potential environmental applications
[J].
Flocculation characterization of a bioflocculant from Bacillus licheniformis
[J].
DMC-grafted cellulose as green-based flocculants for agglomerating fine kaolin particles
[J].
Microwave assisted synthesis and characterization of a novel bio-based flocculant from dextran and chitosan
[J].
Evaluation of the flocculation performance of carboxymethyl chitosan-graft-polyacrylamide,a novel amphoteric chemically bonded composite flocculant
[J].
Implications for industrial application of bioflocculant demand alternatives to conventional media:Waste as a substitute
[J].
A review on chitosan-based flocculants and their applications in water treatment
[J].
A population balance model for flocculation of colloidal suspensions by polymer bridging
[J].
Removal of trace nonylphenol from water in the coexistence of suspended inorganic particles and NOMs by using a cellulose-based flocculant
[J].
Optimization of microbial flocculant-producing medium for Bacillus subtilis
[J].
Production,characterization and structural modification of exopolysaccharide-based bioflocculant by Rhizobium radiobacter SZ4S7S14 and media optimization
[J].
Optimization and characterization of polysaccharide-based bioflocculant produced by Paenibacillus elgii B69 and its application in wastewater treatment
[J].
Production and characterization of a bioflocculant produced by Aspergillus flavus
[J].
Valorization of glycerol/ethanol-rich wastewater to bioflocculants:Recovery,properties,and performance
[J].
Bioflocculant production from untreated corn stover using Cellulosimicrobium cellulans L804 isolate and its application to harvesting microalgae
[J].
Preparation and application of a novel bioflocculant by two strains of Rhizopus sp. using potato starch wastewater as nutrilite
[J].
Production and application of a novel bioflocculant by multiple-microorganism consortia using brewery wastewater as carbon source
[J].
Production of a novel bioflocculant by culture of Klebsiella mobilis using dairy wastewater
[J].
Removal of arsenite by a microbial bioflocculant produced from swine wastewater
[J].
Pretreatment of food waste for methane and hydrogen recovery:A review
[J].
Characterization and application of bioflocculant prepared by Rhodococcus erythropolis using sludge and livestock wastewater as cheap culture media
[J].
Production of bioflocculant using feather waste as nitrogen source and its use in recycling of straw ash-washing wastewater with low-density and high pH property
[J].
Development of a new culture medium for bioflocculant production using chicken viscera
[J].
Bioflocculant production by a consortium of Streptomyces and Cellulomonas species and media optimization via surface response model
[J].
Identification of key genes involved in polysaccharide bioflocculant synthesis in Bacillus licheniformis
[J].
The genome of Xanthomonas campestris pv. campestris B100 and its use for the reconstruction of metabolic pathways involved in xanthan biosynthesis
[J].
Draft genome of the xanthan producer Xanthomonas campestris NRRL B-1459(ATCC 13951)
[J].
Bacterial exopolysaccharides:Chemical structures,gene clusters and genetic engineering
[J].
Towards a better production of bacterial exopolysaccharides by controlling genetic as well as physico-chemical parameters
[J].
Cloning,expression and characterization of a phosphoglucomutase/phosphomannomutase from sphingan-producing Sphingomonas sanxanigenens
[J].
Improvement of exopolysaccharide production in Lactobacillus casei LC2W by overexpression of NADH oxidase gene
[J].
Production of xanthan gum by Sphingomonas bacteria carrying genes from Xanthomonas campestris
[J].
Azotobacter vinelandii lacking the Na+-NQR activity:A potential source for producing alginates with improved properties and at high yield
[J].
Enhancement of transparent hydrogel sanxan production in Sphingomonas sanxanigenens NX02 via rational and random gene manipulation
[J].
Xanthan chain length is modulated by increasing the availability of the polysaccharide copolymerase protein GumC and the outer membrane polysaccharide export protein GumB
[J].
Homologous over-expression of chain length determination protein EpsC increases the molecular weight of exopolysaccharide in Streptococcus thermophilus 05-34
[J].
Identification of substituent groups and related genes involved in salecan biosynthesis in Agrobacterium sp. ZX09
[J].
Recent advances in polysaccharide bio-based flocculants
[J].
Graft copolymeric flocculant using functionalized starch towards treatment of blast furnace effluent
[J].
Plasma-initiated polymerization of chitosan-based CS-g-P(AM-DMDAAC) flocculant for the enhanced flocculation of low-algal-turbidity water
[J].
Flocculation of quaternary chitosan on acid dyeing waste water
[J].
Fabricating an enhanced sterilization chitosan-based flocculants:Synthesis,characterization,evaluation of sterilization and flocculation
[J].
Comparison of two cationic chitosan-based flocculants prepared by photocatalysis and photoinitiation systems:Synthesis mechanism,structure and performance in water treatment
[J].
CSC-P(AM-AA)重金属捕集絮凝剂的光谱表征及其对重金属去除研究
[J].
Spectral characterizations of CSC-P(AM-AA)with function of trapping heavy metals and its removal efficiency of Cu2+
[J].
The role of sulfonated chitosan-based flocculant in the treatment of hematite wastewater containing heavy metals
[J].
Chelating stability of an amphoteric chelating polymer flocculant with Cu(Ⅱ),Pb(Ⅱ),Cd(Ⅱ),and Ni(Ⅱ)
[J].
Flocculation activity and evaluation of chitosan-based flocculant CMCTS-g-P(AM-CA) for heavy metal removal
[J].
Synergistic effect and mechanisms of compound bioflocculant and AlCl3 salts on enhancing Chlorella regularis harvesting
[J].
Natural polymer matrix as safe flocculant to remove turbidity from kaolin suspension:Performance and governing mechanism
[J].
Research on the coagulant aid effects of modified diatomite on coal microbial flocculation
[J].
Synchronous removal of CuO nanoparticles and Cu2+ by polyaluminum chloride-Enteromorpha polysaccharides:Effect of Al species and pH
[J].
N-3-oxo-octanoyl-homoserine lactone as a promotor to improve the microbial flocculant production by an exopolysaccharide bioflocculant-producing bacterium Agrobacterium tumefaciens F2
[J].
The addition of N-hexanoyl-homoserine lactone to improve the microbial flocculant production of Agrobacterium tumefaciens strain F2,an exopolysaccharide bioflocculant-producing bacterium
[J].
The adsorption behavior and mechanism investigation of Pb(Ⅱ) removal by flocculation using microbial flocculant GA1
[J].
Heavy metals removal from wastewater using extracellular polymeric substances produced by Cloacibacterium normanense in wastewater sludge supplemented with crude glycerol and study of extracellular polymeric substances extraction by different methods
[J].
Microbial decolouration of azo dyes:A review
[J].
Textile dye removal from wastewater effluents using bioflocculants produced by indigenous bacterial isolates
[J].
Enhanced composting leachate treatment using extracellular polymeric substances as bioflocculant
[J].
High flocculation of coal washing wastewater using a novel bioflocculant from Isaria cicadae GZU6722
[J].
Application of a modified biological flocculant in total nitrogen treatment of leather wastewater
[J].
Characterisation of fungal bioflocculants and its application in water treatment
[J].
Characterization and flocculability of a novel proteoglycan produced by Talaromyces trachyspermus OU5
[J].


 津公网安备 12010602120337号
津公网安备 12010602120337号{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}